
 English
English
Có 104 OTU được chia sẻ giữa các mẫu HW và DW và 132 OTU được chia sẻ giữa các mẫu HG và DG, lần lượt chiếm 64,20% và 71,74% tổng số (Hình 4a). Các OTU vi sinh vật được chia sẻ giữa ruột tôm khỏe mạnh và nước ao tương ứng chiếm 39,71% tổng số, trong khi tỷ lệ này tăng lên 55,50%, khi tôm bị bệnh.

Hình 4. Phân tích cấu trúc và thành phần vi khuẩn. (a) Phân tích sơ đồ Venn của OTU. (b) Thành phần vi khuẩn ở cấp độ ngành. (c) Thành phần vi khuẩn ở cấp chi. Sự phong phú của vi khuẩn trong cộng đồng tương ứng với giá trị trung bình của mỗi nhóm.
Trong 4 loại mẫu, các đơn vị phân loại vi khuẩn chiếm ưu thế bao gồm Proteobacteria và Bacteroidota ở cấp độ ngành, và sự phong phú của Proteobacteria trong các mẫu khỏe mạnh (nước ao và ruột) cao hơn so với các mẫu bệnh. Actinobacteriota trong các mẫu DW và Firmicutes trong các mẫu DG cũng chiếm ưu thế (Hình 4b). Rhodobacteraceae có sự phong phú cao trong cả 4 loại mẫu (Hình 4c). Algoriphagus rất phong phú trong tất cả các mẫu khỏe mạnh, và Phaeodactylibacter là một trong những đơn vị phân loại vi khuẩn chiếm ưu thế trong tất cả các mẫu nước ao. Ralstonia chiếm ưu thế tuyệt đối (77,75%) trong các mẫu HG, trong khi sự phong phú của Vibrio là cao nhất (50,04%) trong các mẫu DG.
3.3. Phân tích ý nghĩa của sự khác biệt về sự phong phú của vi khuẩn
Kết quả phân tích LEfSe toàn diện cho thấy 16, 5, 9 và 6 dấu ấn sinh học từ ngành đến cấp độ chi lần lượt trong các mẫu HW, HG, DW và DG (Hình 5a). Kết quả phân tích LEfSe riêng biệt của nước ao và mẫu ruột tôm cho thấy thông tin chi tiết hơn về các dấu ấn sinh học khác nhau cho cùng một môi trường sống ở trạng thái khỏe mạnh và bị bệnh (Hình 5b, c). Các dấu ấn sinh học chính trong các mẫu HW chứa chi Algoriphagus, và họ Cyclobacteriaceae, và bộ Cytophagales. Chitinophagales (cấp bộ), Saprospiraceae (cấp họ) và Phaeodactylibacter (cấp chi) là những dấu ấn sinh học chính trong các mẫu DW. Trong ruột tôm khỏe mạnh, các dấu ấn sinh học chính bao gồm Ralstonia (cấp chi), và họ Burkholderiaceae, bộ burkholderiales, lớp Gammaprotebacteria và ngành Protebacteria. Chi Vibrio, và họ Vibrionaceae, và bộ Vibrionales, cũng như Mycoplasmataceae là những dấu ấn sinh học chính trong ruột tôm bị bệnh.

Hình 5. Phân tích sự khác biệt của các đơn vị phân loại trội. (a) Biểu đồ phân cấp loài đa cấp LEfSe cho thấy sự phân bố phát sinh loài của các dòng vi khuẩn trong các môi trường sống khác nhau. (b, c) Phân tích phân biệt tuyến tính cho thấy vi khuẩn chỉ thị trong các môi trường sống khác nhau. Điểm LDA >4.0.
Trong các mẫu HW, sự phong phú tương đối của Ruegeria cao hơn đáng kể, trong khi sự phong phú tương đối của Phaeodactylibacter, Candidatus Aquiluna, Microbacteriaceae và Vibrio thấp hơn đáng kể so với các mẫu DW (P < 0,05, Hình 6a). Sự phong phú tương đối của Ralstonia, Algoriphagus, Burkholderia-Caballeronia-Paraburkholderia và Pelomonas trong các mẫu HG cao hơn đáng kể so với các mẫu DG (P < 0,05, Hình 6b), trong khi sự phong phú tương đối của Vibrio và Paracoccus trong các mẫu DG cao hơn đáng kể (P < 0,05).

Hình 6. Phân tích STAMP về sự khác biệt về sự phong phú của vi khuẩn giữa các mẫu khỏe mạnh và mẫu bệnh bằng cách sử dụng thử nghiệm Welch’s t test. a) Hệ vi sinh vật trong nước ao. (b) Hệ vi sinh vật trong ruột tôm. *P < 0,05, * * P < 0,01, * ** P < 0,001.
3.4. Phân lập và xác định vi khuẩn trội trong đường ruột của tôm bị bệnh
Hầu hết các đàn được phân lập từ tôm bị bệnh đều có đặc điểm tương tự trên đĩa thạch TCBS. Sau nhiều thế hệ lọc, một chủng tinh khiết đã thu được, được đặt tên là TPDS. Kết quả giải trình tự gen cho thấy trình tự 16 S rDNA của chủng TPDS (GenBank số gia nhập No. ON566023) là 1509 bp (Bảng bổ sung S2). So sánh tương đồng trình tự được thực hiện trong cơ sở dữ liệu RDPII và thu được 20 chủng loại liên quan chặt chẽ (Bảng bổ sung S3), tất cả đều là Vibrio. Phân tích cây phát sinh loài cho thấy chủng TPDS có liên quan chặt chẽ nhất với V. owensii và V. hyugaensis (Hình 7).
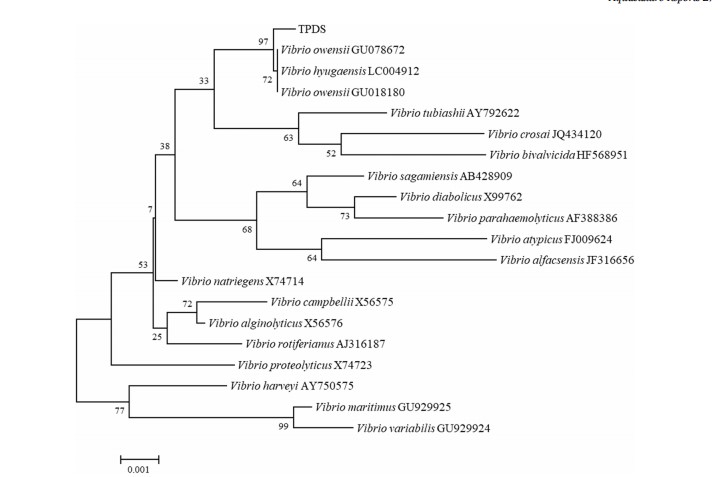
Hình 7. Cây phát sinh chủng loài của TPDS dựa trên trình tự gen 16 S rDNA một phần sử dụng phân tích NJ. Phần trăm giá trị bootstrap (1000 bản sao).
3.5. Dự đoán chức năng của cộng đồng vi khuẩn đường ruột
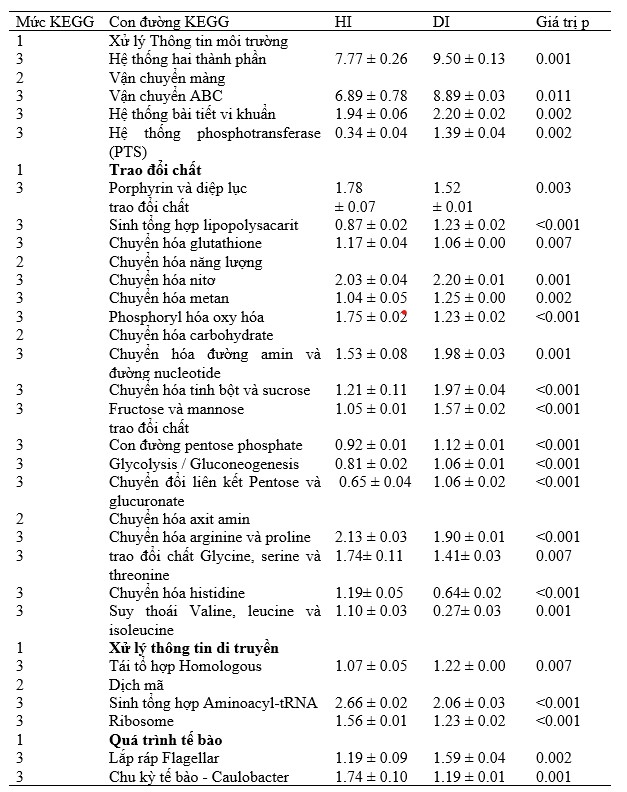
Dựa trên các dự đoán chức năng metagenomic, các gen liên quan đến chuyển hóa carbohydrate (chuyển hóa đường amin và đường nucleotide, chuyển hóa tinh bột và sucrose, chuyển hóa fructose và mannose, con đường pentose phosphate, glycolysis / gluconeogenesis và chuyển đổi pentose và glucuronate), sinh tổng hợp và chuyển hóa glycan (sinh tổng hợp lipopolysacarit), vận chuyển màng (chất vận chuyển ABC, hệ thống bài tiết vi khuẩn, hệ thống phosphotransferase), truyền tín hiệu (hai thành phần hệ thống) và khả năng vận động của tế bào (lắp ráp flagellar) đã được làm giàu đáng kể trong các mẫu DG (P < 0,01, Hình 8; Bảng 1). Ngược lại, sự phong phú tương đối của các gen liên quan đến chuyển hóa axit amin (chuyển hóa arginine và proline, chuyển hóa glycine, serine và threonine, chuyển hóa histidine, thoái hóa valine, leucine và isoleucine), chuyển hóa các đồng yếu tố và vitamin (chuyển hóa porphyrin và diệp lục), chuyển hóa năng lượng (phosphoryl hóa oxy hóa), chuyển hóa lipid, sao chép và sửa chữa, dịch mã (sinh tổng hợp aminoacyl-tRNA, ribosome), gấp lại, phân loại và thoái hóa, và tế bào tăng trưởng và chết (chu kỳ tế bào Caulobacter) cao hơn đáng kể trong các mẫu HG (P < 0,05).

Hình 8. Phân tích sự phong phú tương đối của các chức năng dự đoán ở KEGG cấp 2 bằng cách sử dụng thử nghiệm Welch’s t test. * P < 0,05, * * P < 0,01, * ** P < 0,001.
Bảng 1. Sự phong phú tương đối (%) của các hàm dự đoán. KEGG cấp 1 và cấp độ 2, cũng như cấp độ 3 được liệt kê.
 4. Thảo luận
4. Thảo luận
Một căn bệnh mới trên PL Penaeus vannamei, được gọi là BVS hoặc TPD, đã bùng phát ở Trung Quốc trong những năm gần đây, gây thiệt hại kinh tế rất lớn. Các nghiên cứu đã chỉ ra rằng PL tôm mắc bệnh thường có các triệu chứng bao gồm hoại tử và bong tróc tế bào biểu mô của ống gan tụy và ruột (Zou và cộng sự, 2020; Wang và cộng sự, 2021a, 2021b), cũng được tìm thấy trong nghiên cứu này. Những bằng chứng này cho thấy gan tụy và ruột tôm PL là cơ quan mục tiêu chính của mầm bệnh. Tuy nhiên, chưa có báo cáo nào tập trung vào mối quan hệ giữa các cộng đồng vi khuẩn môi trường hoặc đường ruột và căn bệnh này hiện nay. Trong nghiên cứu này, chúng tôi đã phát hiện ra sự khác biệt về sự đa dạng, sự phong phú tương đối và chức năng của hệ vi sinh vật môi trường và đường tiêu hóa giữa tôm khỏe mạnh và bị bệnh.
Đặc biệt, hệ vi sinh vật trong ruột, đóng vai trò quan trọng trong việc duy trì sức khỏe của vật chủ (Rooks và Garrett, 2016). Nói chung, có một mối quan hệ giữa mất đa dạng vi khuẩn đường ruột và bệnh tật, cũng được chứng minh ở nhiều loại sinh vật, bao gồm cả con người (Clarke và cộng sự, 2014). Các nghiên cứu trước đây đã được báo cáo rằng sự xuất hiện của bệnh “mang đỏ” ở cá và “WFS” ở tôm đi kèm với sự đa dạng vi khuẩn đường ruột bị tổn thương (Li và cộng sự, 2017; Hou và cộng sự, 2018). Tuy nhiên, trong một số nghiên cứu, hiện tượng xuất hiện bệnh tôm và sự gia tăng đa dạng vi khuẩn đường ruột đã được quan sát đồng thời (Cornejo-Granados và cộng sự, 2017; Zhou và cộng sự, 2019). Trong nghiên cứu của chúng tôi, các chỉ số của Shannon và Smithwilson đã tăng đáng kể, và chỉ số Simpson đã giảm đáng kể trong ruột tôm bị bệnh, điều này cho thấy sự đồng đều và đa dạng trong ruột tôm bị bệnh cao hơn so với ở những cá thể khỏe mạnh. Do đó, cần có thêm bằng chứng và nghiên cứu chuyên sâu hơn để chứng minh mối quan hệ giữa bệnh và sự đa dạng của hệ vi sinh vật đường ruột.
Đáng chú ý, bằng cách so sánh ruột tôm khỏe mạnh với ở nước ao, chúng tôi thấy rằng sự đa dạng và độ đồng nhất của nước ao cao hơn đáng kể. PCA và phân tích cụm cho thấy các cộng đồng vi sinh vật trong các môi trường sống khác nhau được tách biệt rõ ràng và số lượng OTU được chia sẻ tương đối thấp giữa chúng. Những bằng chứng này cho thấy hệ vi sinh vật đường ruột của tôm khỏe mạnh PL ít bị ảnh hưởng bởi hệ vi sinh vật nước. Tương tự, các nghiên cứu trước đây đã chứng minh rằng môi trường sống là yếu tố chính chiếm ưu thế trong các đặc điểm cộng đồng vi khuẩn (Xiong và cộng sự, 2018, 2019; Zeng và cộng sự, 2020). Điều thú vị là sự khác biệt về chỉ số Simpson vi khuẩn của nước ao và ruột trong các mẫu bệnh trở nên không còn đáng kể, khoảng cách phân cụm của cấu trúc cộng đồng vi khuẩn trong hai đã được rút ngắn và số lượng OTU được chia sẻ tăng lên. Những kết quả này cho thấy hệ vi sinh vật đường ruột của tôm bị bệnh bị ảnh hưởng đáng kể bởi hệ vi sinh vật nước. Sự xuất hiện của bệnh đi kèm với sự thay đổi trong quá trình lắp ráp của cộng đồng vi khuẩn đường ruột (Dini-Andreote và cộng sự, 2015; Zhu và cộng sự, 2016). Trong khi tầm quan trọng của các quá trình xác định giảm dần, thì các quá trình ngẫu nhiên chiếm ưu thế trong việc thúc đẩy tập hợp cộng đồng, cho thấy rằng sự xuất hiện của bệnh sẽ làm giảm tác động lọc của vật chủ đối với các loài ngoại lai (Xiong và cộng sự, 2016, 2018; Yao và cộng sự, 2018). Do đó, đặc điểm của cộng đồng vi khuẩn trong ruột tôm bị bệnh cho thấy nhiều điểm tương đồng hơn với trong nước ao, đã được tìm thấy trong nghiên cứu về các bệnh khác của tôm (Liang và cộng sự, 2020).
Proteobacteria và Bacteroidota là những vi khuẩn chiếm ưu thế nhất trong môi trường nuôi trồng thủy sản (Chen và cộng sự, 2019), cũng đã được chứng minh trong nghiên cứu này. Khi bệnh khởi phát, sự phong phú của Phaeodactylibacter, Candidatus Aquiluna, Microbacteriaceae và Vibrio đã tăng lên đáng kể, trong khi sự phong phú của Ruegeria đã giảm đáng kể, cho thấy thành phần cộng đồng vi khuẩn bình thường trong nước ao đã bị xáo trộn. Các nghiên cứu đã chỉ ra rằng Proteobacteria có nhiều nhất trong ruột tôm (Holt và cộng sự, 2021). Đáng chú ý, sự phong phú của Proteobacteria trong ruột tôm khỏe mạnh cao hơn đáng kể so với tôm bị bệnh, trong đó sự phong phú của Ralstonia là nhiều nhất. Ralstonia thường được cho là vi khuẩn gây bệnh trên thực vật (Xue và cộng sự, 2020; Fluit và cộng sự, 2021), và một số thậm chí đã được phát hiện là có hại cho con người (Carreira và cộng sự, 2020). Tuy nhiên, trong lĩnh vực nuôi trồng thủy sản, Ralstonia đã được quan sát là vi khuẩn cốt lõi trong ruột của một số loài cá, có khả năng sản xuất enzyme cao (Wu và cộng sự, 2021). Ngoài ra, một số nghiên cứu đã chỉ ra rằng Ralstonia có tiềm năng trở thành chế phẩm sinh học cho nuôi trồng thủy sản (Van Hung, Bossier và cộng sự, 2019; Văn Hùng và cộng sự, 2019). Quan trọng hơn, sự tích tụ đáng kể của Mycoplasmataceae và Vibrio đã được quan sát thấy trong ruột tôm bị bệnh. Một số loài Mycoplasmataceae thường được coi là mầm bệnh phổ biến gây bệnh cho người và động vật (Leal Zimmer và cộng sự, 2020), nhưng khả năng gây bệnh của chúng trong nuôi trồng thủy sản hiện vẫn đang gây tranh cãi (Cheaib và cộng sự, 2021). Một số nghiên cứu chỉ ra rằng Vibrio đã gây bệnh nặng ở tôm (Yu và cộng sự, 2021; Aguilar-Rend’on và cộng sự, 2022). Trong nghiên cứu của chúng tôi, Vibrio sp. chiếm ưu thế được phân lập từ ruột tôm được xác định là gần nhất với V. owensii và V. hyugaensis, và biểu hiện cao của chúng có thể là nguyên nhân trực tiếp gây bệnh.
Những thay đổi trong cấu trúc của hệ vi sinh vật đường ruột chắc chắn dẫn đến sự khác biệt về chức năng gen của chúng. Trong nghiên cứu này, sự khác biệt về chức năng cộng đồng vi khuẩn giữa tôm khỏe mạnh và tôm bị bệnh chủ yếu được phản ánh trong quá trình trao đổi chất và xử lý thông tin môi trường. Dựa trên cơ sở dữ liệu con đường KEGG, sự phong phú của các gen liên quan đến chuyển hóa carbohydrate trong ruột tôm bị bệnh đã tăng lên đáng kể, trong khi sự phong phú của các gen liên quan đến giảm, cho thấy sự xuất hiện của bệnh đã thay đổi chế độ trao đổi chất của hệ vi sinh vật đường ruột. Trong một nghiên cứu về Vibrio cholerae, chuyển hóa carbohydrate đã được chứng minh là cung cấp nguồn năng lượng chính cho sự tăng trưởng, sản xuất yếu tố độc lực và xâm chiếm trong vật chủ (Van Alst và DiRita, 2020). Do đó, sự gia tăng các con đường trao đổi chất carbohydrate có thể tạo điều kiện thuận lợi vật chủ nhiễm Vibrio. Các con đường KEGG liên quan đến xử lý thông tin môi trường đã tăng đáng kể ở ruột tôm bị bệnh, bao gồm hệ phản ứng hai thành phần và vận chuyển màng (chất vận chuyển ABC, hệ thống bài tiết vi khuẩn và hệ thống phosphotransferase), cả hai đều đã được chứng minh là điều chỉnh sự bài tiết và vận chuyển xuyên màng của các yếu tố gây bệnh (Green và Mecsas, 2016; De Silva và Kumar, 2019; Rom và cộng sự, 2021; Zeng và Charkowski, 2021). Ngoài ra, các gen liên quan đến tái tổ hợp tương đồng và lắp ráp flagellar cũng được quan sát thấy được kích hoạt trong ruột của tôm bị bệnh, có liên quan chặt chẽ đến khả năng gây bệnh của vi khuẩn. Tuy nhiên, chức năng của các gen này chỉ được dự đoán trong nghiên cứu này và vẫn cần được chứng thực bằng nghiên cứu sâu hơn với nhiều cách tiếp cận hơn.
Tóm lại, nghiên cứu này cho thấy mối quan hệ tương tác giữa Bệnh mờ đục trên tôm post và hệ vi sinh vật của nước ao hoặc ruột tôm. Sự phong phú của Mycoplasmataceae và Vibrio, và sự ức chế của Ralstonia trong ruột tôm có thể là lý do quan trọng cho sự xuất hiện của bệnh này. Trong khi đó, bệnh có thể làm thay đổi quá trình lắp ráp của hệ vi sinh vật đường ruột tôm, khiến nó dễ bị ảnh hưởng bởi hệ vi sinh vật trong nước, do đó tiếp tục gây ra sự xáo trộn về sự đa dạng, thành phần và chức năng của hệ vi sinh vật đường ruột. Quan trọng nhất, nghiên cứu này có thể giúp hiểu sâu hơn về nguyên nhân, phòng ngừa và kiểm soát bệnh này trong quá trình nuôi tôm giống.
Theo Peng Yu, Hongwei Shan, Yu Cheng, Jingjing Ma, Kai Wang, Hongyang Li
Nguồn: https://www.sciencedirect.com/science/article/pii/S2352513422004069
Biên dịch: Đoàn Thị Huyền Thoại – Nguyễn Thị Quyên
TÔM GIỐNG GIA HÓA – CHÌA KHÓA THÀNH CÔNG
Xem thêm:
- Vấn Đề Dịch Bệnh Lớn Nhất Đối Với Nghề Nuôi Tôm Hiện Nay Là Gì? AHPNS?
- Rà Soát Lại Bệnh Vibriosis Trong Nuôi Tôm
- Bột Và Dầu Ruồi Lính Đen Làm Nguyên Liệu Sản Xuất Thức Ăn Chức Năng Cho Tôm Biển