
 English
English
Tóm tắt
Giới thiệu: Ngành nuôi tôm thẻ chân trắng Litopenaeus vannamei phát triển nhanh chóng phần lớn phụ thuộc vào việc cung ứng được ấu trùng chất lượng cao. Sự phát triển của ấu trùng tôm là một quá trình biến thái độc đáo, nhưng quá trình này cũng có thể làm tôm chết do dễ nhiễm bệnh. Hiện nay, các nghiên cứu về miễn dịch bẩm sinh ở tôm chủ yếu tập trung vào giai đoạn trưởng thành mà ít nghiên cứu về miễn dịch bẩm sinh ở ấu trùng.
Phương pháp: Trong nghiên cứu này, sử dụng phương pháp giải trình tự Illumina thông lượng cao, chúng tôi so sánh bản phiên mã của ấu trùng L. vannamei Zoea và những con trưởng thành bị nhiễm Vibrio parahaemolyticus.
Kết quả: Tổng cộng 487.887.650 lượt đọc chất lượng cao đã được thu thập, tập hợp và tổng hợp thành 23.802 gen, trong đó 3315 được xác định là các gen biểu hiện khác nhau. Sau đó, phân tích làm giàu giới hạn GO, phân tích làm giàu con đường KEGG và phân tích gen chức năng đã được thực hiện. Chúng tôi nhận thấy rằng các con đường liên quan đến phản ứng miễn dịch bẩm sinh, chẳng hạn như con đường truyền tín hiệu mTOR, con đường truyền tín hiệu MAPK và con đường truyền tín hiệu Notch, đã phong phú hơn ở giai đoạn Zoea so với giai đoạn trưởng thành, cho thấy khả năng miễn dịch bẩm sinh chống lại Vibrio vulnificus có thể tồn tại trong giai đoạn ấu trùng Zoea.
Thảo luận: Ở giai đoạn Zoea phong phú hơn so với giai đoạn trưởng thành, cho thấy khả năng miễn dịch bẩm sinh chống lại Vibrio vulnificus có thể tồn tại trong giai đoạn ấu trùng Zoea. Những phát hiện này sẽ cung cấp thông tin hữu ích cho việc nghiên cứu khả năng miễn dịch bẩm sinh ở ấu trùng tôm.
1. Giới thiệu
Tôm thẻ chân trắng Litopeneaus vannamei, là một trong những loài nuôi trồng thủy sản phổ biến nhất trên toàn thế giới, đạt sản lượng 4.966.200 tấn (FAO, 2020). Việc nuôi tôm thẻ chân trắng bị cản trở chủ yếu do chất lượng ấu trùng kém và dịch bệnh bùng phát thường xuyên (Wang Y. và cộng sự, 2020). Ương dưỡng ấu trùng đề cập đến các giai đoạn phát triển của ấu trùng tôm, bao gồm giai đoạn Nauplius, Zoea, Mysis và giai đoạn đầu tôm post (Hertzler và Freas, 2009). Đó là một quá trình quan trọng quyết định phần lớn sự thành công của nuôi tôm thẻ chân trắng L. vannamei. Trong các giai đoạn nuôi tôm tiếp theo, sự sinh trưởng, phát triển và khả năng kháng stress/bệnh của tôm tỷ lệ thuận với chất lượng của ấu trùng (Angthong và cộng sự, 2021).
Trong giai đoạn đầu phát triển của tôm, hình thái, sinh lý và sinh thái của chúng trải qua những thay đổi sâu sắc. Ở giai đoạn ấu trùng, đặc biệt là từ Zoea đến tôm post, tôm có khả năng miễn dịch thấp và dễ bị nhiễm nhiều mầm bệnh, có thể dẫn đến tôm chết hàng loạt trong trại giống (Zheng và cộng sự, 2016). Vibrio Parahemolyticus là vi khuẩn gram âm có khả năng gây bệnh mạnh và khả năng lây truyền cao cho động vật thủy sản (Velázquez-Lizárraga và cộng sự, 2019). Nhiễm Vibrio parahaemolyticus ở giai đoạn đầu của tôm, đặc biệt là giai đoạn Zoea, thường khiến số lượng lớn ấu trùng chết (Angthong và cộng sự, 2021). Gần đây, một căn bệnh đặc biệt được gọi là “bệnh mờ đục trên ấu trùng tôm” (TPD) hoặc “bệnh hậu ấu trùng thủy tinh” (GPD) do Vibrio có độc lực cao gây ra, đã gây ra thiệt hại nghiêm trọng về mặt kinh tế ở Trung Quốc (Zou và cộng sự, 2020).
Tương tự như các động vật không xương sống khác, tôm phụ thuộc vào khả năng miễn dịch bẩm sinh bao gồm các phản ứng của thể dịch và tế bào để chống lại nhiễm trùng. Sự điều hòa của hệ thống prophenoloxidase (ProPO), protein đông máu, melanine và peptide kháng khuẩn đều là thành phần của phản ứng miễn dịch dịch thể (Mycology và cộng sự, 1998). Phản ứng miễn dịch tế bào loại bỏ mầm bệnh bằng các quá trình tạo máu như thực bào, apoptosis, hình thành nốt sần và bao bọc (Jiravanichpaisal và cộng sự, 2006). Tuy nhiên, phần lớn các phản ứng miễn dịch này được phát hiện ở giai đoạn trưởng thành của L. vannamei và có rất ít thông tin về hệ thống miễn dịch bẩm sinh trong giai đoạn phát triển ban đầu của L. vannamei. Nghiên cứu này đã phân tích bản phiên mã của ấu trùng Zoea và những con trưởng thành L. vannamei bị nhiễm Vibrio parahaemolyticus bằng kỹ thuật giải trình tự Illumina thông lượng cao. Để khám phá các gen liên quan đến miễn dịch và các quá trình sinh học liên quan đến phản ứng kháng mầm bệnh ở giai đoạn ấu trùng Zoea, dữ liệu phiên mã ở giai đoạn trưởng thành được so sánh với dữ liệu phiên mã giai đoạn ấu trùng. Nghiên cứu này sẽ cung cấp những hiểu biết sâu sắc về phản ứng miễn dịch bẩm sinh ở ấu trùng Zoea của tôm thẻ chân trắng L. vannamei đối với các bệnh nhiễm trùng do vi khuẩn gây bệnh.
2. Vật liệu và phương pháp
2.1. Tôm và mầm bệnh
Giai đoạn ấu trùng (giai đoạn 4–5) và tôm trưởng thành (trọng lượng trung bình 10,55 ± 1,97 g) của tôm thẻ chân trắng L. vannamei được thu thập từ trang trại nuôi tôm Hengxing ở thành phố Trạm Giang, Quảng Đông, Trung Quốc. Không yêu cầu giấy phép đặc biệt đối với các địa điểm và hoạt động lấy mẫu, đồng thời các nghiên cứu không bao gồm các loài và địa điểm có nguy cơ tuyệt chủng hoặc được bảo vệ.
Ấu trùng được thuần, rửa kỹ bằng nước biển, khử trùng bằng Iốt (100 μl/ L) và duy trì trong bể thủy tinh 200 L ở 30–31°C với nước biển được lọc 0,45 μm. Từ giai đoạn Zoea 1, chúng được cho ăn hỗn hợp Isochrisis galbana (30%) và Chaetoceros muelleri (70%) với mật độ tối thiểu 75.000 tế bào/ ml. Nước được thay 30% mỗi ngày và ấu trùng được nuôi trong quang kỳ 12 giờ. Trước mỗi thí nghiệm, ấu trùng được quan sát bằng kính hiển vi để đánh giá hoạt động, sự biến dạng, tình trạng túi noãn hoàng, ký sinh trùng và sự bám dính của mảnh vụn.
Tôm được thuần hoá trong một tuần ở phòng thí nghiệm. Nước trong bể nuôi được duy trì ở nhiệt độ 26 ± 0,5°C, độ mặn 3‰, pH 7,2 ± 0,2 và thay 1/3 sau mỗi 48h. Sục khí được cung cấp để duy trì oxy hòa tan ở nồng độ khoảng 8 mg/L. Tôm được cho ăn thức ăn công thức với tỷ lệ 1% trọng lượng cơ thể vào lúc 8 giờ sáng và 5 giờ chiều mỗi ngày.
Vibrio parahaemolyticus được lấy từ Trung tâm Thu thập Vi sinh Quảng Đông, GDMCC số 1.306. Vibrio parahaemolyticus được cấy qua đêm vào môi trường Luria–Bertani broth ở 37°C và khuấy ở tốc độ 150 vòng/phút. Các khuẩn lạc được thu thập và tạo huyền phù trong nước muối vô trùng thông thường (NaCl 0,9%) và ly tâm ở tốc độ 6000×g trong 5 phút ở 20°C. Điều chỉnh huyền phù vi khuẩn đến mật độ quang học là 1,0 ở bước sóng 610 nm và được pha loãng theo thứ tự để đạt được mật độ 10,5 CFU/ ml.
2.2. Nhiễm Vibrio parahaemolyticus và lấy mẫu
Thí nghiệm ấu trùng Zoea cảm nhiễm Vibrio parahaemolyticus được chia thành nhóm đối chứng (LC) và nhóm nhiễm Vibrio parahaemolyticus (LE). 100 ấu trùng Zoea được cho cảm nhiễm lần lượt với 1 ml Vibrio parahaemolyticus nồng độ 10,4 cfu/ ml trong 24 giờ và 1 ml PBS. Không thay nước trong suốt thí nghiệm, mỗi nghiệm thức được lặp lại năm lần. 5 ml vi tảo được bổ sung sau mỗi 6 giờ. Sau khi cho ấu trùng ăn vi tảo, dung dịch vi khuẩn đã được tiêm 1 lần. Sử dụng kính hiển vi soi nổi, ấu trùng sống sót được đếm sau 24 giờ. Vào lúc 0 và 24 giờ, các mẫu ấu trùng còn sống sót từ nhóm đối chứng và nhóm thử nghiệm được bảo quản ngay trong nitơ lỏng và sau đó được bảo quản ở -80°C.
Tôm (trọng lượng trung bình 10,55 ± 1,97 g, n=50) được chia thành hai nhóm, nhóm đối chứng (SC) và nhóm cảm nhiễm Vibrio parahaemolyticus (SE). Mỗi nhóm có bốn bể với 50 con tôm trong 200 L nước có sục khí, khử trùng bằng tia cực tím. Theo cách tương tự, Vibrio parahaemolyticus được nuôi cấy với nồng độ vi khuẩn cuối cùng là 2 x 103 CFU/ mL. Tiêm vào cơ tôm 50ul dung dịch vi khuẩn và PBS. Gan tụy của tôm còn sống ở nhóm đối chứng và nhóm thử nghiệm được lấy trong 24 giờ và mẫu ngay lập tức được ngâm trong nitơ lỏng và sau đó được bảo quản ở -80°C.
2.3. Phân lập RNA và giải trình tự Illumina
Tổng RNA được chiết xuất từ mỗi mẫu (100 ấu trùng; 6 con tôm) bằng thuốc thử Trizol (Invitrogen, CA, USA) theo quy trình của nhà sản xuất. Bioanalyzer 2100 và RNA 1000 Nano LabChip Kit (Agilent, CA, USA) đã được sử dụng để phân tích số lượng và độ tinh khiết của tổng số RNA có số RIN > 7,0. RNA poly(A) được tinh chế khỏi tổng RNA (5ug) bằng cách sử dụng các hạt từ tính gắn oligo-T poly-T bằng hai vòng tinh chế. Sau khi tinh chế, mRNA bị phân mảnh thành các mảnh nhỏ sử dụng cation hóa trị hai ở nhiệt độ cao. Sau đó, các đoạn RNA đã cắt được sao chép ngược để tạo dữ liệu cDNA cuối cùng theo phương pháp bộ chuẩn bị mẫu mRNASeq (Illumina, San Diego, Hoa Kỳ), kích thước chèn trung bình cho các dữ liệu đầu cặp là 300 bp (± 50 bp). Cuối cùng, chúng tôi đã thực hiện giải trình tự kết thúc ghép đôi 2×150 bp (PE150) trên Illumina Novaseq™ 6000 (LC-Bio Technology CO., Ltd., Hàng Châu, Trung Quốc) theo phương pháp được đề xuất của nhà cung cấp.
2.4. Lắp ráp và chú thích bản phiên mã de novo
Đầu tiên, Cutadapt (Kechin và cộng sự, 2017) và các lệnh Perl nội bộ được sử dụng để loại bỏ các lần đọc có chứa ô nhiễm bộ điều hợp, các trình tự chất lượng thấp và các trình tự không xác định. Sau đó, chất lượng trình tự đã được xác minh bằng FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/), bao gồm nội dung Q20, Q30 và GC của dữ liệu sạch. Tất cả các phân tích tiếp theo đều dựa trên dữ liệu sạch có chất lượng cao. Việc lắp ráp bản phiên mã De novo được thực hiện với Trinity 2.4.0 (Grabherr và cộng sự, 2011). Nhóm Trinity phiên mã thành các cụm dựa trên nội dung trình tự được chia sẻ. Cụm phiên mã như vậy được gọi là một ‘gen’. Bản phiên mã dài nhất trong cụm được chọn làm chuỗi ‘gen’ (Unigene).
Tất cả các Unigenes được lắp ráp đều được căn chỉnh dựa trên cơ sở dữ liệu protein không dư thừa (Nr) (http://www.ncbi.nlm.nih.gov/), bản thể học gen (GO) (http://www.geneontology.org), SwissProt (http://www.Exasy.ch/sprot/), Bách khoa toàn thư về gen và bộ gen ở Kyoto (KEGG) (http://www.genome.jp/kegg/) và eggnog (http://eggnogdb.embl.de /) cơ sở dữ liệu sử dụng DIAMOND (Buchfink và cộng sự, 2014) với ngưỡng đánh giá <0,00001.
2.5. Phân tích các gen biểu hiện khác nhau (DEG)
Salmon (Patro và cộng sự, 2017) đã sử dụng để thực hiện mức biểu hiện cho Unigenes bằng cách tính toán bảng phiên mã trên một triệu (TPM) (Mortazavi và cộng sự, 2008). Các Unigenes được biểu thị khác nhau đã được chọn có ý nghĩa thống kê (giá trị p < 0,05) bởi gói R edgeR (Robinson và cộng sự, 2009). Tiếp theo,thực hiện phân tích làm giàu GO và KEGG một lần nữa trên Unigenes được thể hiện khác nhau bằng các tập lệnh perl nội bộ.
2.6. Xác thực dữ liệu RNA-seq và khảo sát biểu hiện gen bằng Q-PCR
Phân tích QPCR được thực hiện để xác nhận các mẫu biểu hiện gen thu được từ RNA-seq. Q-PCR được tiến hành với Bộ SYBR® PrimeScript™ RT-PCR (TaKaRa, Trung Quốc) theo phương pháp của nhà sản xuất. Mỗi phản ứng chứa 1× SYBR Premix Ex Taq, 10 μM mỗi mồi và 2 μL cDNA (50 ng/ mL) trong thể tích cuối cùng là 25 μL. Độ tinh khiết và nồng độ của các mẫu cDNA được xác định bằng máy quang phổ kế NanoDrop (ND-8000). 11 gen liên quan đến miễn dịch đã được chọn để xác nhận qPCR. Đoạn mồi cụ thể của từng gen được thiết kế bằng Chương trình Primer Premier (Bảng S2). Mỗi phản ứng qPCR chứa 100 ng mẫu cDNA, 0,2 µM mỗi mồi và 1X SYBR Green SsoAdvanced (BioRad). Các thông số chu kỳ như sau; biến tính ban đầu ở 95°C trong 30 giây để kích hoạt DNA polymerase, sau đó là 40 chu kỳ 5 giây ở 95°C, 30 giây ở 55°C và 30 giây ở 72°C. Cấu hình biểu hiện của từng gen được tính toán bằng phương pháp 2−ΔΔCT (Livak và Schmittgen, 2001). Phân tích biểu hiện gen tương đối đã được chuẩn hóa thành gen tham chiếu (-actin) như một biện pháp kiểm soát nội bộ. Tất cả các qPCR được thực hiện trong ba lần sao chép sinh học (n = 3).
3. Kết quả
3.1. Đặc điểm hình thái của ấu trùng Zoea và tôm trưởng thành nhiễm Vibrio
Sau quá trình biến thái của ấu trùng Nauplius thành ấu trùng Zoea, cơ thể dường như được chia thành nhiều đoạn và miệng mở để kiếm ăn. Ấu trùng Zoea bình thường có lớp giáp đầu ngực trong suốt và các phần phụ có thể di chuyển được và không bám vào vật thể (Hình 1A). Sau 24 giờ nhiễm Vibrio parahaemolyticus, một lượng lớn chất màu đen được quan sát thấy ở vùng giáp đầu ngực của ấu trùng Zoea so với ấu trùng khỏe mạnh. Ấu trùng giảm hoạt động, khả năng ăn kém và cơ thể ấu trùng trở nên dính (Hình 1B). So với tôm bình thường, tôm trưởng thành bị nhiễm Vibrio parahaemolyticus có biểu hiện gan tụy bị teo và ruột không rõ ràng (Hình 1C, D).
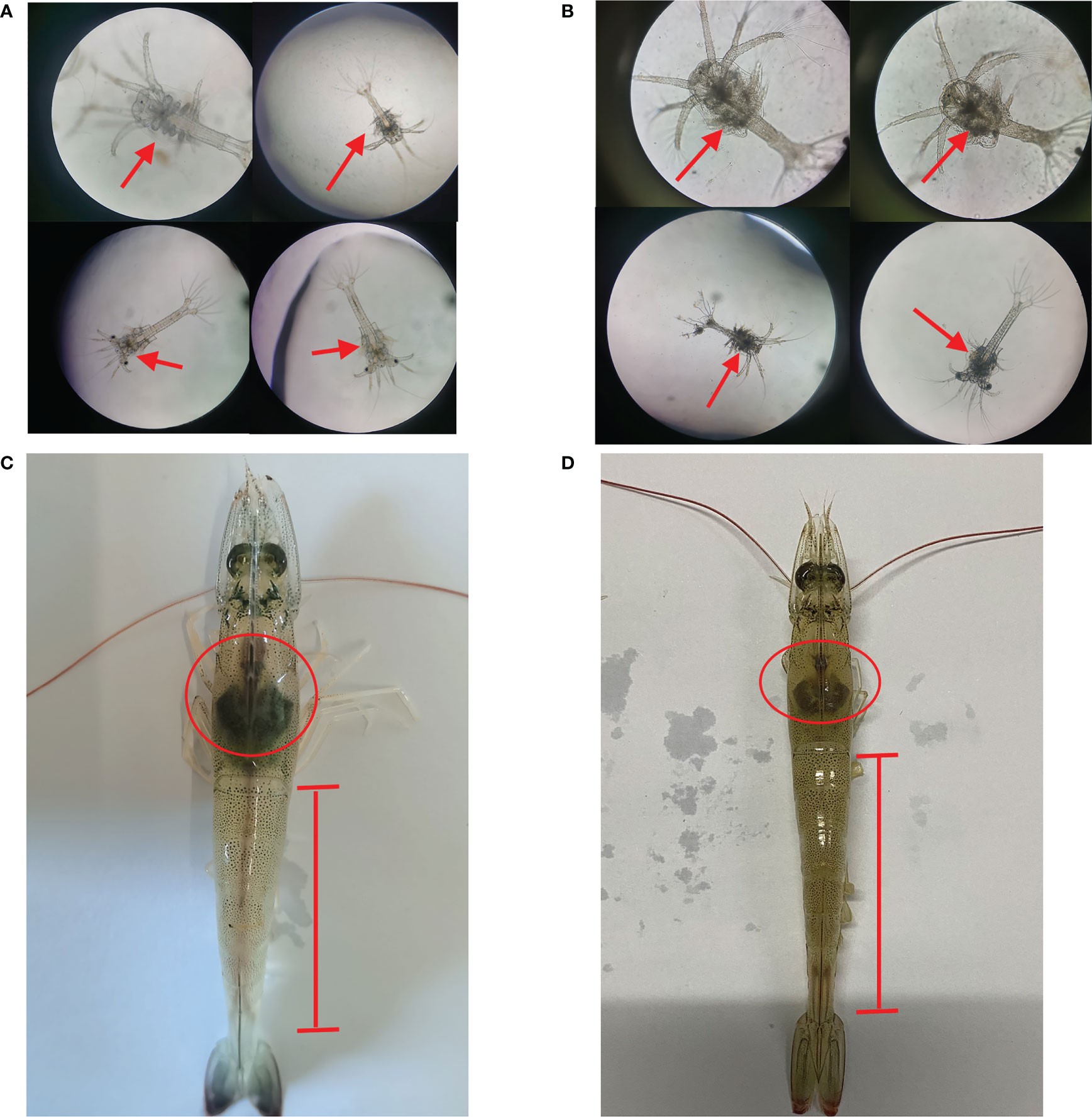
Hình 1. Dấu hiệu bên ngoài của ấu trùng Zoea và tôm trưởng thành bị nhiễm Vibrio parahaemolyticus. (A) Ấu trùng Zoea bình thường. (B) Ấu trùng bị nhiễm Vibrio parahaemolyticus. (C) Tôm trưởng thành bình thường. (D) Tôm trưởng thành bị nhiễm Vibrio parahaemolyticus.
3.2. Lọc dữ liệu và phát hiện DEG sau khi xử lý mầm bệnh
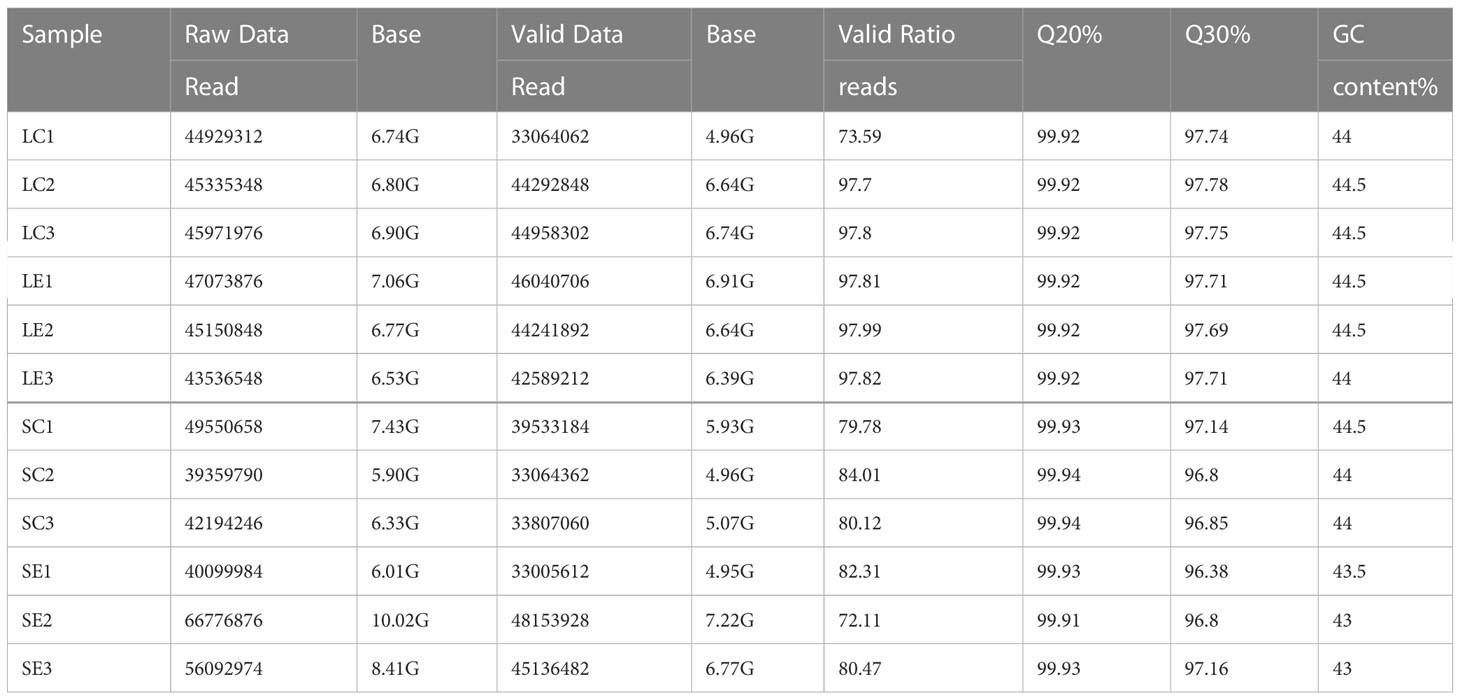
Bằng cách sử dụng máy giải trình tự Illumina Novaseq™ 6000, các mẫu ấu trùng và tôm trưởng thành từ cả nhóm đối chứng và nhóm nhiễm vi khuẩn Vibrio parahaemolyticus đều được xử lý trình tự phiên mã thông lượng cao. Trong mỗi dữ liệu, có khoảng 39.359.790 đến 66.776.776 bài đọc thô được tạo ra. Sau khi loại bỏ các lượt đọc có chứa bộ điều hợp hoặc poly-N và các lượt đọc chất lượng thấp khỏi dữ liệu thô, 33.005.612 đến 48.153.928 lượt đọc hợp lệ đã được lọc ra (Bảng 1). Chất lượng dữ liệu được đảm bảo với Q20 (xác suất lỗi trình tự cơ sở < 1%) > 99% và Q30 (xác suất lỗi trình tự cơ sở < 0,1%) > 96% trong mỗi dữ liệu (Bảng 1).
Bảng 1. Dữ liệu RNA-seq của các mẫu.
Để xác định đặc điểm biểu hiện gen của ấu trùng Zoea và tôm trưởng thành sau khi nhiễm mầm bệnh, bốn dữ liệu gen biểu hiện khác nhau (DEG) đã được xây dựng từ nhóm ấu trùng đối chứng (LC), đối chứng trưởng thành (SC), ấu trùng thí nghiệm (LE) và thí nghiệm trưởng thành (SE). DEG được xác định bằng cách sử dụng phương pháp Bayesian sau khi mức độ biểu hiện gen được chuẩn hóa để đọc trên mỗi kilobase của trình tự phiên mã trên một triệu lần đọc được ánh xạ (RPKM). Bằng cách so sánh các dữ liệu phiên mã của nhóm bị nhiễm mầm bệnh và nhóm đối chứng, tổng số 3.315 gen biểu hiện khác nhau (DEG) đã được phát hiện. Có 1156 gen điều hòa tăng và 578 gen điều hòa giảm trong nhóm LC-LE. Nhóm SC-SE có tổng cộng 1245 gen điều hòa tăng và 336 gen điều hòa giảm (Hình 2A). Nghiên cứu sơ đồ Venn của DEG cho các nhóm khác nhau cho thấy 81 DEG chồng chéo giữa các nhóm LC-LE và SC-SE, nhưng 1653 DEG trong nhóm LC-LE khác biệt với 1500 DEG trong nhóm SC-SE (Hình 2B). 36 DEG được điều chỉnh được chồng chéo giữa các nhóm LC-LE và SC-SE, trong khi 1209 DEG được điều chỉnh trong SC-SE khác biệt với 1120 DEG được điều chỉnh trong LC-LE (Hình 2C). 3 DEG được điều hòa ở mức chồng chéo giữa các nhóm LC-LE và SC-SE, trong khi 575 DEG được điều hòa ở LC-LE khác với 333 DEG được điều hòa ở SC-SE (Hình 2D).
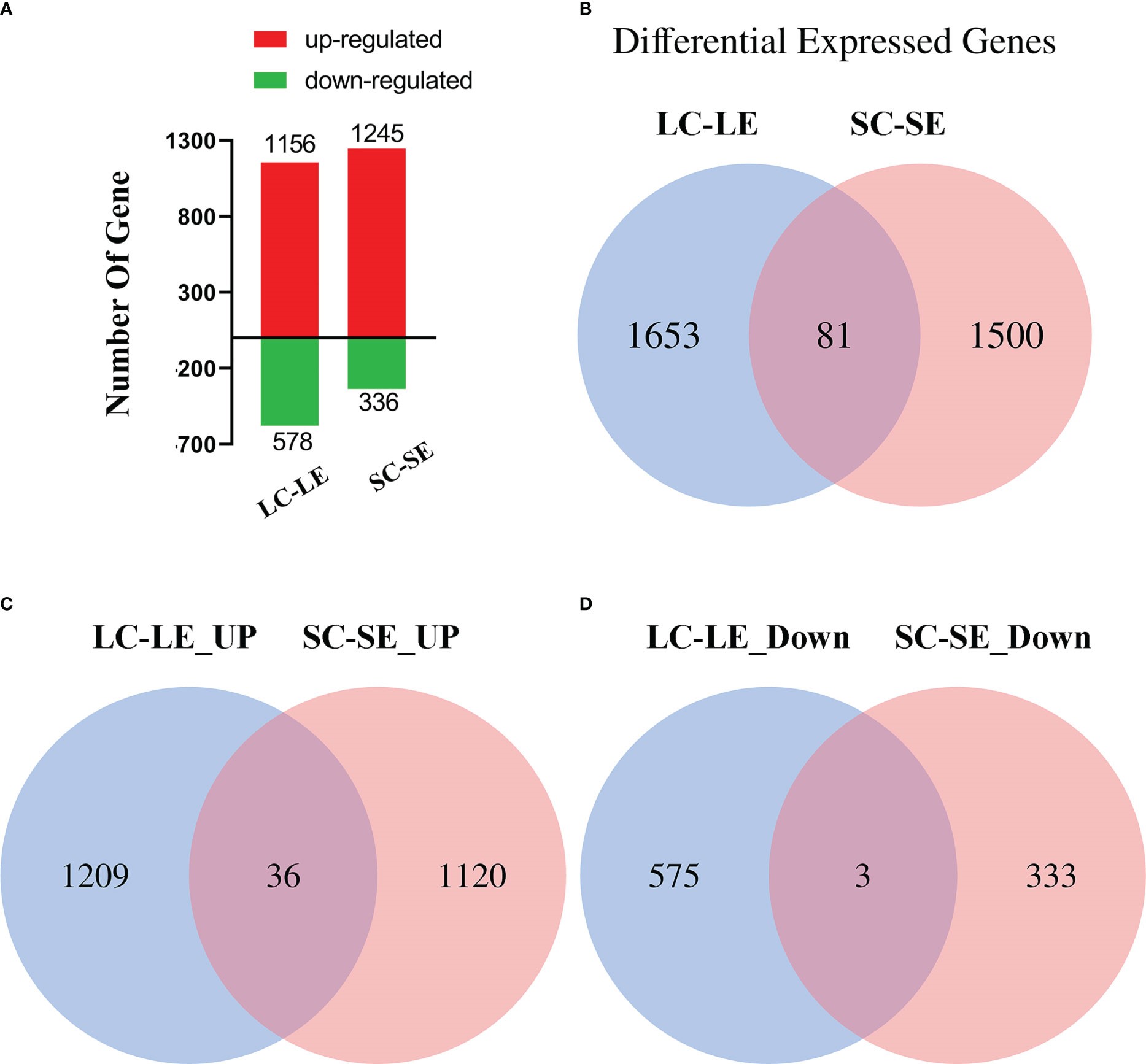
Hình 2. Kết quả thống kê của các gen biểu hiện khác nhau (DEG). (A) Số lượng gen làm việc biểu hiện khác nhau, LC-LE và SC-SE. Thanh màu đỏ cho các gen điều hòa tăng, thanh màu xanh lá cây cho các gen điều hòa giảm. (B) Biểu đồ Venn hiển thị biểu thức 2 lần của bảng phiên mã trong bảng phiên mã LC-LE và SC-SE. (C) Biểu đồ Venn hiển thị biểu thức được điều chỉnh tăng của bảng điểm trong bảng phiên mã LC-LE và SC-SE. (D) Biểu đồ Venn hiển thị biểu thức được điều chỉnh giảm của bảng phiên mã trong bảng phiên mã LC-LE và SC-SE. Kết quả phân tích thống kê đều có Padj < 0,05.
3.3. Chú thích chức năng của DEG
Để đánh giá thêm chức năng của DEG và các đường truyền tín hiệu mà chúng tham gia, 3315 DEG đã được phân loại thành các loại quá trình sinh học (BP), chức năng phân tử (MF) và thành phần tế bào (CC) (Hình 3A, B). Trong số các bản phiên mã của nhóm LC-LE, các bản phiên mã hiển thị các thuật ngữ phong phú sau đây về quá trình sinh học: phân giải protein (46 bản phiên mã), quá trình trao đổi chất chitin (30 bản phiên mã) và quá trình phosphoryl hóa protein (27 bản phiên mã). Đối với chức năng phân tử, các thuật ngữ được làm phong phú là: liên kết protein (47 bản phiên mã), liên kết ATP (45 bản phiên mã) và liên kết ion kim loại (39 bản phiên mã). Thành phần không thể thiếu của màng (122 bản phiên mã), tế bào chất (114 bản phiên mã) và màng sinh chất (93 bản phiên mã) là từ khóa GO cho các thành phần tế bào (Hình 3A). Trong nhóm SC-SE, các thuật ngữ dành cho bản phiên mã cho thấy các thuật ngữ sau đây dành cho quá trình sinh học: phân giải protein (61 bản phiên mã), quá trình oxy hóa-khử (37 bản phiên mã) và bám dính tế bào (25 bản phiên mã); đối với chức năng phân tử: liên kết với protein (50 bản phiên mã), liên kết ion kim loại (48 bản phiên mã) và liên kết ion canxi (48 bản phiên mã); đối với thành phần tế bào: tế bào chất (139 bản phiên mã), thành phần không thể thiếu của màng (124 bản phiên mã) và màng sinh chất (105 bản phiên mã) (Hình 3B).
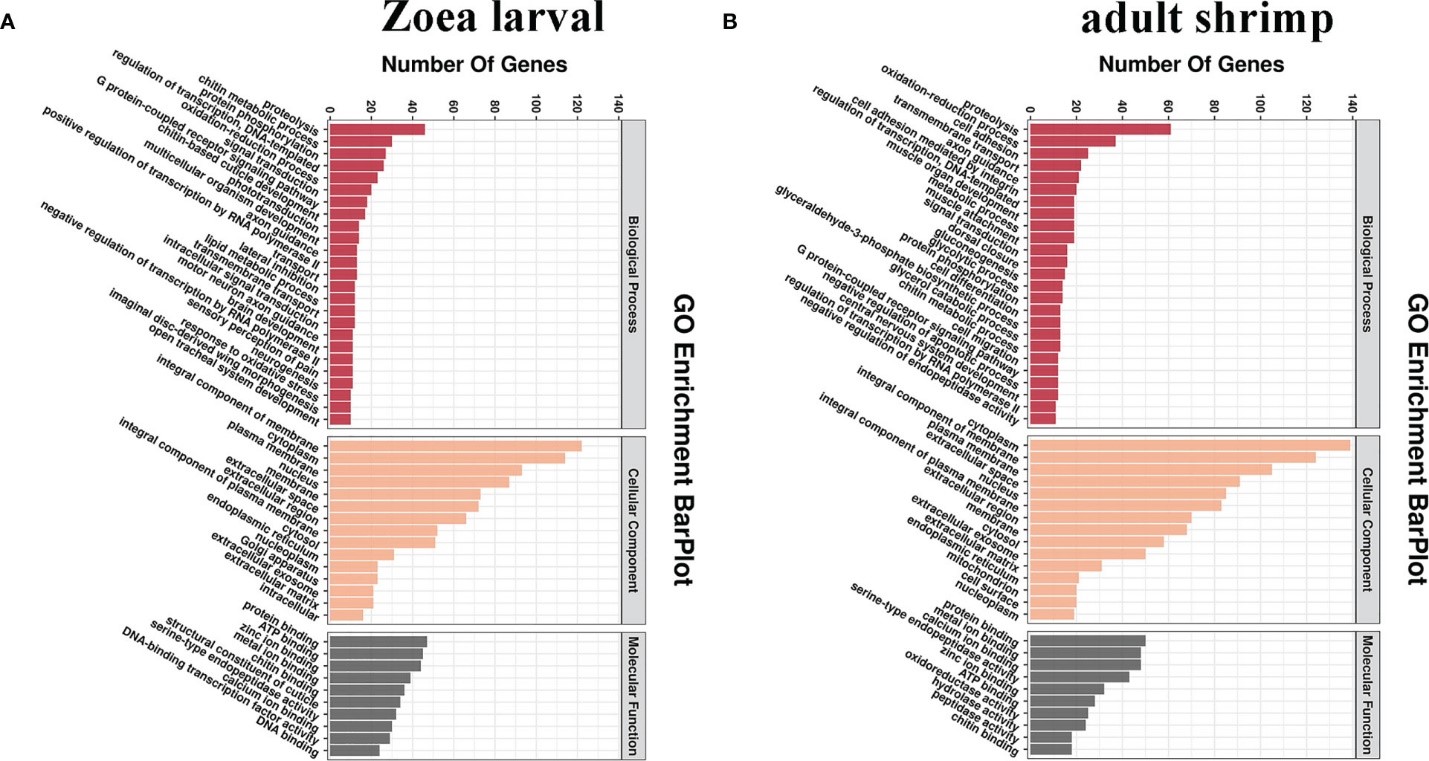
Hình 3. Phân công bản thể gen cho các gen biểu hiện khác nhau (DEG) ở ấu trùng Zoea và tôm trưởng thành sau khi nhiễm V. parahaemolyticus. Kết quả phân loại GO được tóm tắt thành ba loại GO chính (thành phần tế bào, chức năng phân tử và quá trình sinh học). (A) Phân tích GO để tìm các gen trên ấu trùng Zoea bị nhiễm V. parahaemolyticus. (B) Phân tích GO cho các gen trên tôm trưởng thành bị nhiễm V. parahaemolyticus.
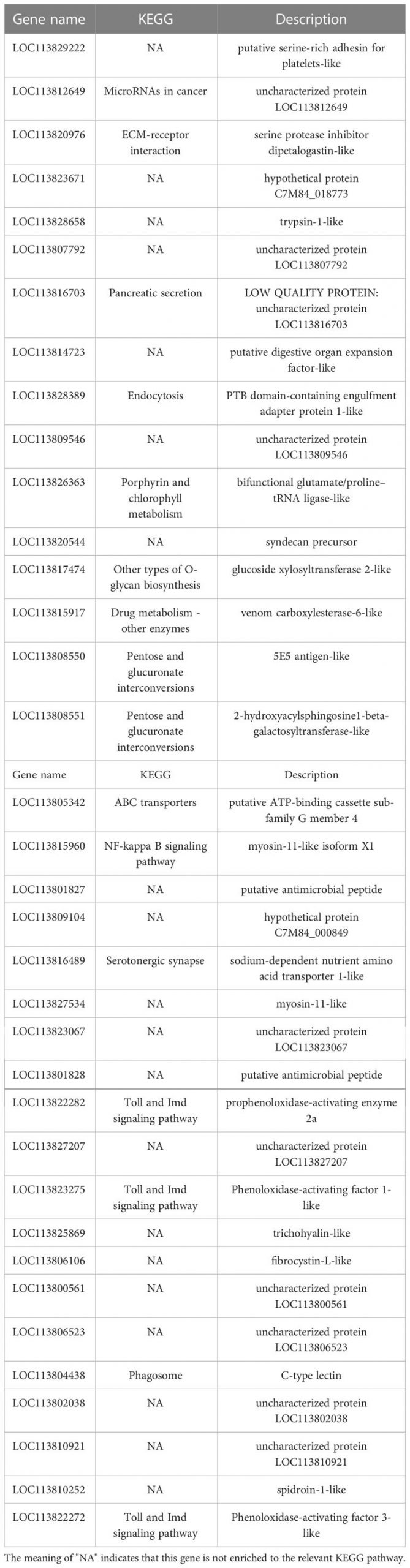
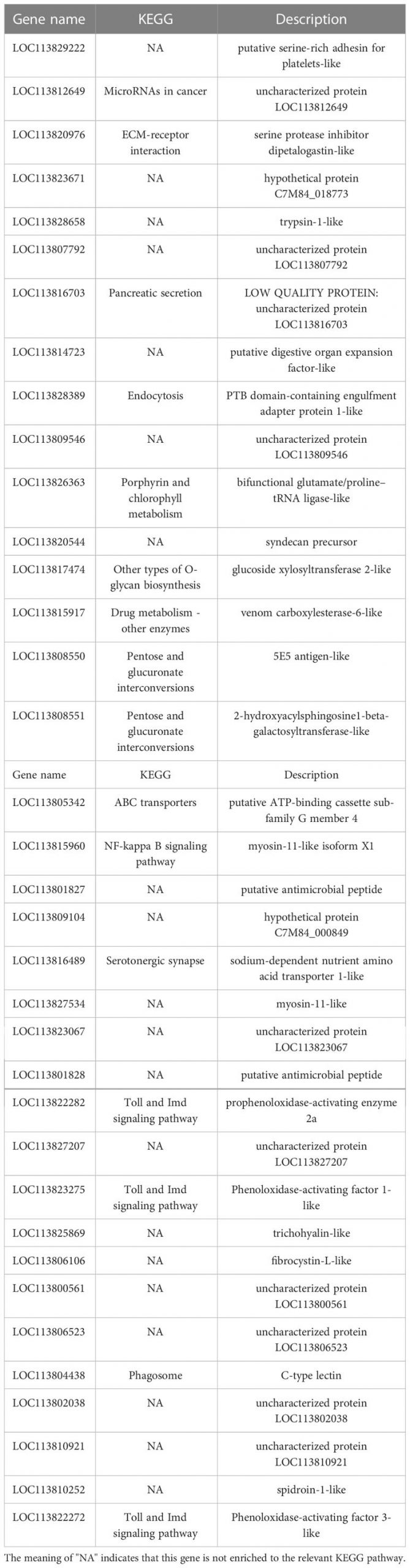
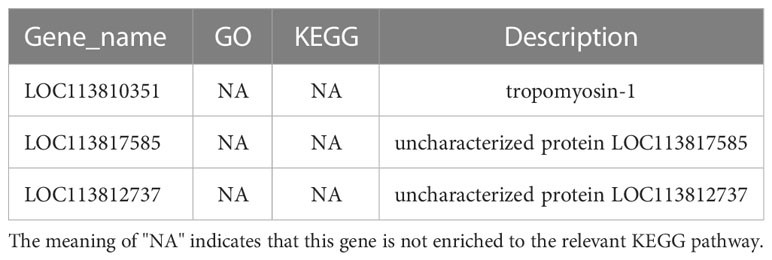
Cơ sở dữ liệu con đường KEGG là cơ sở dữ liệu để phân tích sản phẩm gen trong quá trình trao đổi chất và các chức năng gen liên quan trong quá trình tế bào. Phân tích dựa trên con đường giúp chúng ta hiểu rõ hơn về chức năng sinh học của gen (Yi và cộng sự, 2020). 551 DEG được tìm thấy trong các nhóm LC-LE được liên kết với 236 con đường KEGG đã biết trong cuộc điều tra này. 402 DEG được liên kết với 178 con đường KEGG đã biết cho các nhóm SC-SE. Trên cơ sở một nghiên cứu trước đó về sơ đồ Venn của DEG, các phân tích làm giàu KEGG cho cả DEG được điều chỉnh tăng và giảm trong mỗi nhóm đã được tiến hành (Hình 4A–D). Các DEG được điều chỉnh đặc biệt trong nhóm LC-LE chủ yếu được làm phong phú theo các con đường miễn dịch như đường dẫn tín hiệu MAPK, các tầng bổ sung và đông máu cũng như đường dẫn tín hiệu mTOR (Hình 4A). Việc điều chỉnh tăng các DEG dành riêng cho nhóm SC-SE, chủ yếu được làm phong phú thêm trong các con đường miễn dịch bao gồm tương tác với thụ thể ECM, đường dẫn tín hiệu Lysosome, Toll và Imd, đường dẫn tín hiệu VEGF và đường dẫn tín hiệu MAPK (Hình 4B). Các DEG được điều hòa giảm đặc biệt trong nhóm LC-LE tập trung vào quá trình dẫn truyền quang – ruồi, chuyển hóa Purine và con đường chuyển hóa Fructose và mannose (Hình 4C). Các DEG được điều chỉnh giảm cụ thể trong nhóm SC-SE chủ yếu tập trung vào quá trình chuyển hóa Lysosome, Arginine và proline cũng như các con đường chuyển đổi xen kẽ Pentose và glucuronate (Hình 4D). Tổng cộng có 36 DEG được điều hòa trong nhóm LC-LE và SC-SE (Bảng 2), trong đó các gen liên quan đến miễn dịch là: chất ức chế serine protease giống dipetalogastin, lectin loại C, yếu tố kích hoạt Phenoloxidase 3 giống, Yếu tố kích hoạt phenoloxidase giống 1, enzyme kích hoạt prophenoloxidase 2a và peptide kháng khuẩn giả định. Các con đường miễn dịch được làm phong phú ở cả hai gen được điều hòa lên là: tương tác với thụ thể ECM, Endocytosis, đường dẫn tín hiệu NF-kappa B, đường dẫn tín hiệu Toll và Imd và phagosome. Ba gen được điều hòa giảm phổ biến trong nhóm LC-LE và SC-SE (Bảng 3), tropomyosin-1, protein không bị biến đổi LOC113817585 và protein không bị biến đổi LOC113812737.
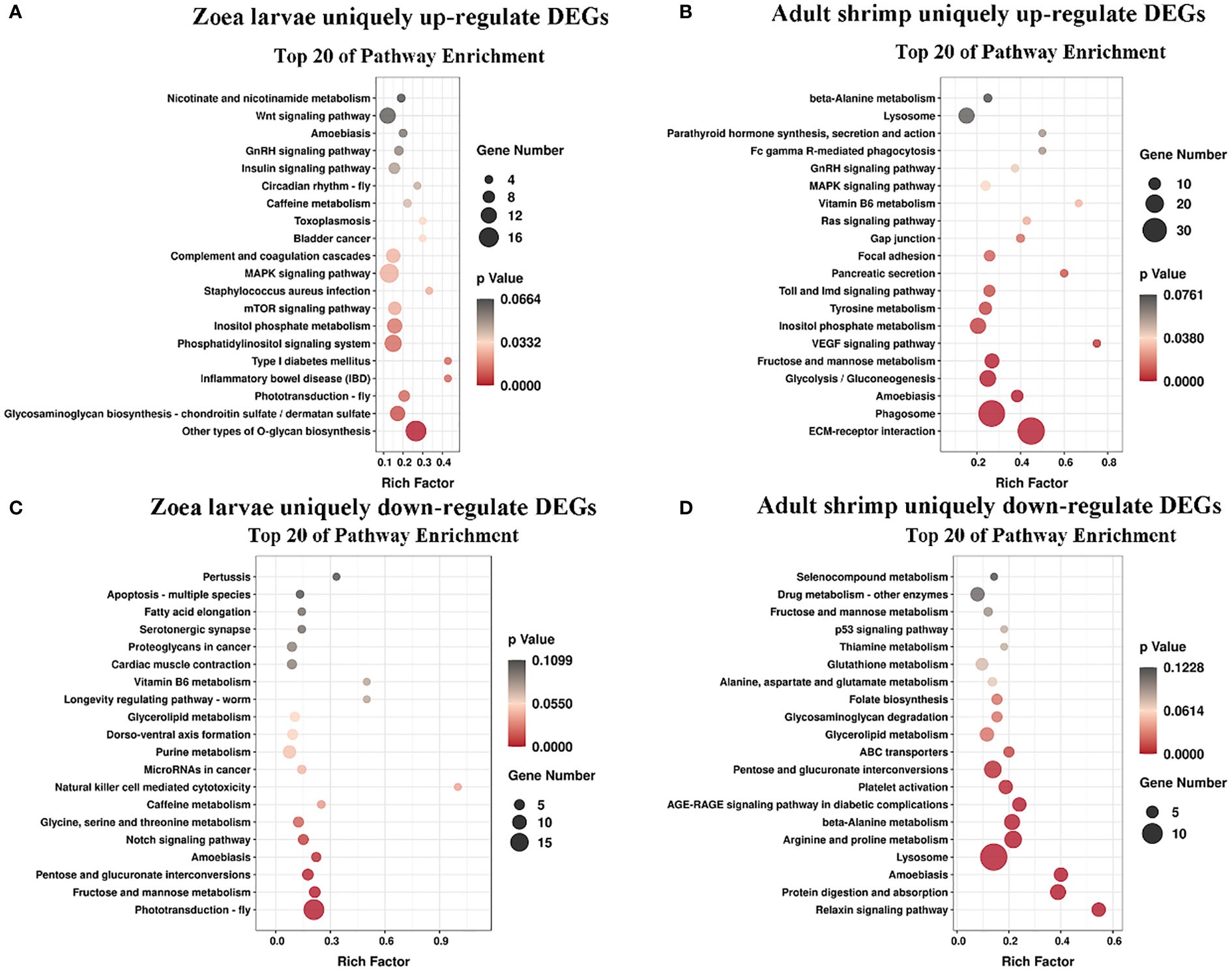
Hình 4 Sơ đồ phân tán làm giàu con đường KEGG. Trục biểu thị tên của đường dẫn, trục hoành biểu thị hệ số phong phú tương ứng của đường dẫn, kích thước của giá trị p được biểu thị bằng màu của điểm. Số lượng gen khác biệt được biểu thị bằng kích thước của điểm. (A) Làm phong phú thêm các thuật ngữ KEGG bằng các gen điều chỉnh tăng duy nhất sau khi ấu trùng Zoea bị nhiễm Vibrio parahaemolyticus. (B) Làm phong phú thuật ngữ KEGG của các gen điều chỉnh tăng duy nhất sau khi nhiễm Vibrio parahaemolyticus ở con trưởng thành. (C) Làm phong phú thêm các thuật ngữ KEGG bằng các gen điều hòa giảm duy nhất sau khi ấu trùng Zoea bị nhiễm Vibrio parahaemolyticus. (D) Làm phong phú thêm các thuật ngữ KEGG dành riêng cho các gen bị điều hòa giảm sau khi nhiễm Vibrio parahaemolyticus ở con trưởng thành.
Bảng 2. Sự lây nhiễm của ấu trùng Zoea và con trưởng thành với Vibrio parahaemolyticus đồng điều hòa 36 gen.
Bảng 3. Sự nhiễm khuẩn của ấu trùng Zoea và con trưởng thành với 3 gen đồng điều hòa vi khuẩn Vibrio parahaemolyticus.
3.4. Xác thực các mẫu biểu hiện của DEG bằng qPCR
Để xác thực các kiểu biểu hiện của DEG sau khi bị nhiễm Vibrio parahaemolyticus ở ấu trùng Zoea và tôm trưởng thành, chúng tôi đã chọn 8 DEG để phân tích qRT-PCR (Hình 5) bao gồm 5 gen điều hòa tăng và 3 gen điều hòa giảm. Cấu hình biểu thức của tất cả các IRD được chọn từ qPCR được so sánh với cấu hình biểu thức từ RNA-seq. Những thay đổi mạnh mẽ về mức độ biểu hiện gen của 8 DEG này sau khi kích thích trong 24 giờ: β-Arrestin1, Protein yippee-like 1, tiền chất protein gắn với beta-1,3-glucan, giống yếu tố kháng lipopolysaccharide và kích hoạt phenoloxidase giống yếu tố 3 được điều chỉnh tăng đáng kể, trong khi mức độ biểu hiện của isoform X1 giống adenosylhomocysteinase, giống glutathione peroxidase và giống trypsin-1 được điều chỉnh giảm đáng kể.
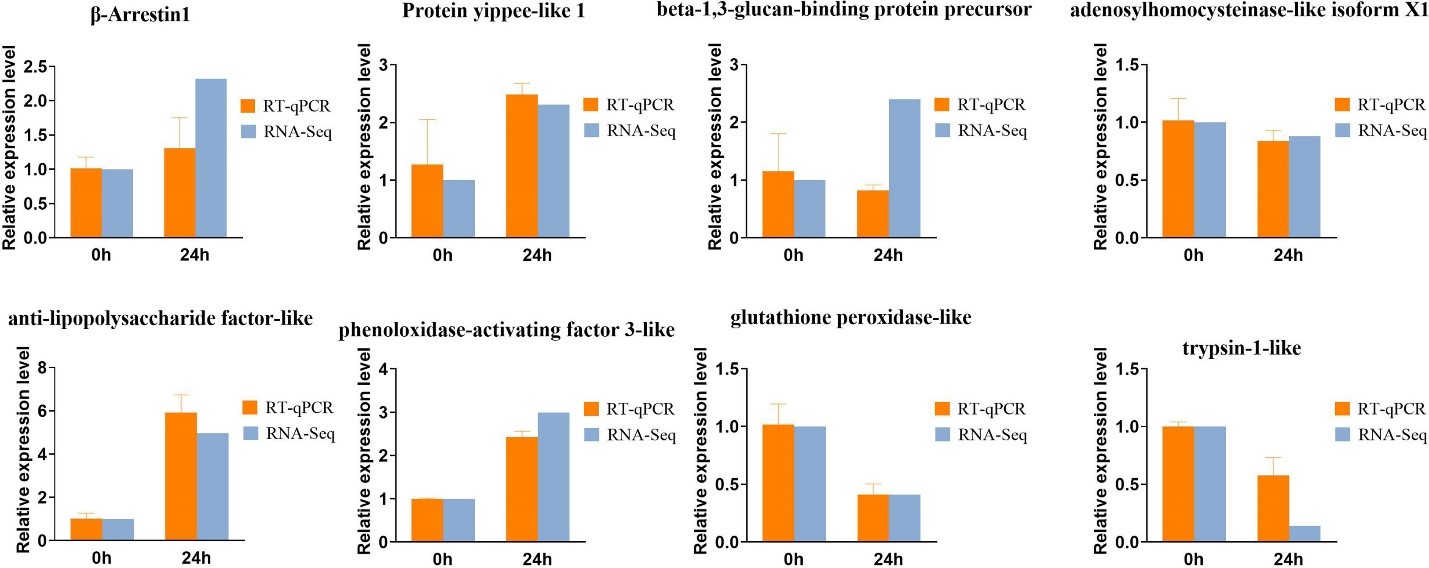
Hình 5. Các bản phiên mã từ dữ liệu RNAseq đã được xác minh bằng phương pháp PCR thời gian thực định lượng (qPCR), trong đó các gen liên quan đến ấu trùng Zoea là beta-Arrestin1, Protein yippee-like 1, beta-1, tiền chất protein gắn 3-glucan, adenosine homocysteinase -like isoform X1, và trên người trưởng thành có chất giống yếu tố kháng lipopolysaccharide, chất kích hoạt 3 giống phenol oxydase, giống glutathione peroxidase và giống trypsin. Các thanh lỗi cho biết lỗi tiêu chuẩn của giá trị trung bình từ ba lần sinh học. Các chữ cái khác nhau thể hiện sự khác biệt có ý nghĩa theo ANOVA (giá trị p < 0,05).
4. Thảo luận
Là một thành viên của nhóm giáp xác, tôm thẻ chân trắng (Litopenaeus vannamei) trải qua một số phát triển biến thái trong giai đoạn đầu (Wei và cộng sự, 2014). Chế độ phát triển ban đầu độc đáo này dẫn đến những thay đổi mạnh mẽ về cả mô hình dinh dưỡng và cấu trúc hình thái thông qua quá trình biến thái và chúng dễ mắc bệnh hơn do hệ thống miễn dịch kém phát triển. Đặc biệt, chế độ dinh dưỡng thay đổi đáng kể từ dự trữ lòng đỏ sang dinh dưỡng săn mồi ở ấu trùng đến Zoea (Aguirre-Guzmán và cộng sự, 2001). Trong thời kỳ ấu trùng Zoea, ấu trùng sống dựa vào nguồn thức ăn bên ngoài và nhạy cảm hơn với các tác động từ môi trường bên ngoài, đặc biệt là nhiễm vi khuẩn Vibrio gây bệnh cản trở sự phát triển của ấu trùng. Có báo cáo cho rằng phân tích trình tự phiên mã đã được thực hiện trong suốt giai đoạn phát triển ban đầu của tôm với mục đích phân tích so sánh những thay đổi sinh lý trong quá trình biến thái của tôm (Wei và cộng sự, 2014 ; Angthong và cộng sự, 2021). Hiện nay, phần lớn các nghiên cứu tập trung vào giai đoạn trưởng thành, ít nghiên cứu điều tra khả năng miễn dịch chống Vibrio ở giai đoạn đầu của tôm.
Trong nghiên cứu này, chúng tôi thực hiện phân tích phiên mã của tôm thẻ Litopenaeus vannamei bị nhiễm Vibrio parahaemolyticus ở cả giai đoạn ấu trùng và trưởng thành. Trong nghiên cứu giải trình tự, toàn bộ ấu trùng được sử dụng, trong khi mô gan tụy được lấy từ con trưởng thành. Gan tụy là cơ quan chính để điều trị dự phòng miễn dịch ở tôm (Ji và cộng sự, 2009), và vì ấu trùng quá nhỏ để lấy mẫu gan tụy ấu trùng nên ấu trùng phải được thu thập toàn bộ. Chú thích GO của unigen được phân thành ba loại chức năng: quá trình sinh học, thành phần tế bào và chức năng phân tử (Hình 3B). Các thuật ngữ GO được thể hiện quá mức cho quá trình sinh học là quá trình phân giải protein và quá trình oxy hóa-khử. Trong danh mục thành phần tế bào, ba thuật ngữ GO đã được làm phong phú đáng kể: tế bào chất, thành phần không thể thiếu của màng và màng sinh chất. Liên kết protein và liên kết ion kim loại là các chức năng phân tử phổ biến nhất. Việc kích hoạt phản ứng miễn dịch của tôm phụ thuộc vào nhiều cơ chế khác nhau, bao gồm hệ thống chống oxy hóa, tan máu, chữa lành vết thương và các protein liên quan đến Apoptosis (Tassanakajon và cộng sự, 2013). Trong quá trình trao đổi chất hiếu khí bình thường, tất cả các sinh vật đều tạo ra các loại oxy phản ứng (ROS). Nhiễm vi khuẩn trong tình trạng căng thẳng có thể dẫn đến thiếu oxy và tăng ROS, dẫn đến căng thẳng oxy hóa nội bào (Livingstone, 2001). Mức độ ROS tăng lên gây ra tổn thương đại phân tử, từ đó ảnh hưởng đến cấu trúc màng và enzyme cũng như tính toàn vẹn của axit nucleic (Indo và cộng sự, 2007). Sau khi trưởng thành bị nhiễm Vibrio parahaemolyticus, các gen được điều chỉnh lên xuống khác nhau này đã được kiểm tra độc lập để làm giàu theo con đường KEGG (Hình 4B). Các con đường giàu KEGG như Toll (Huang và cộng sự, 2010; Yang và cộng sự, 2007), suy giảm miễn dịch (Imd) (Lan và cộng sự, 2013; Wang và cộng sự, 2019), con đường truyền tín hiệu MAPK (Yan và cộng sự., 2013) và con đường phagosome (Liu và cộng sự, 2020) đã được chứng minh là có liên quan đến phản ứng miễn dịch của tôm đối với các mầm bệnh xâm nhập. Điều này phù hợp với những phát hiện về con đường làm giàu gen miễn dịch ở tôm trưởng thành bị nhiễm Vibrio parahaemolyticus trong nghiên cứu này. Con đường và các gen được điều hòa nhiều nhất ở tôm nhiễm Vibrio là con đường tương tác với thụ thể ECM. Ở động vật giáp xác, một số báo cáo cho thấy con đường tương tác giữa thụ thể ECM có thể liên quan đến phản ứng miễn dịch chống lại nhiễm trùng do vi khuẩn (Yang và cộng sự, 2018), nhiễm virus hội chứng đốm trắng (WSSV) (Zhong và cộng sự, 2017) và phơi nhiễm kim loại nặng (Meng và cộng sự, 2015). Con đường truyền tín hiệu của yếu tố tăng trưởng nội mô mạch máu (VEGF) được biết là đóng một vai trò quan trọng trong sự tăng sinh tế bào nội mô, di cư, hình thành mạch, tính thấm của mạch máu, ức chế apoptosis và nhiễm virus (Li và cộng sự, 2017; Wang và cộng sự, 2019). Trong nghiên cứu này, các gen được điều hòa tăng cường đã được làm phong phú theo con đường này và có thể con đường này có liên quan đến khả năng miễn dịch chống Vibrio.
Nhiễm Vibrios là một trong những nguyên nhân chính gây chết ở giai đoạn đầu của tôm. Trong nghiên cứu này, Vibrio parahaemolyticus được chọn để lây nhiễm ấu trùng Zoea và tôm trưởng thành. Mục đích của chúng tôi là xác định bất kỳ gen và con đường giống hệt hoặc khác biệt nào phản ứng với sự lây nhiễm Vibrios của ấu trùng hoặc tôm trưởng thành. Cụm từ quy trình sinh học GO quan trọng nhất đối với nhóm LC-LE là “phân giải protein”, tiếp theo là “quá trình trao đổi chất chitin” và “quá trình phosphoryl hóa protein”. Trong danh mục thành phần tế bào, ba thuật ngữ GO thành phần không thể thiếu của màng, tế bào chất và màng sinh chất đã được làm phong phú đáng kể. Liên kết với protein, liên kết ATP và liên kết ion kim loại là các chức năng phân tử phổ biến nhất (Hình 3A). Việc nhiễm ấu trùng Zoea với Vibrio parahaemolyticus tạo ra ATP thông qua các phản ứng oxy hóa của chính cơ thể, thúc đẩy quá trình tổng hợp và thoái hóa protein, đồng thời kích hoạt quá trình phosphoryl hóa protein và các hoạt động khác để chống lại sự xâm nhập của mầm bệnh (Tang và cộng sự, 2022). Ở động vật có vú, chitin được phát hiện có tác dụng kích hoạt các tế bào miễn dịch bẩm sinh và gây viêm đường hô hấp ở chuột (Koller và cộng sự, 2011). Nó cũng là thành phần chính của bộ xương ngoài của cua và tôm, và hoạt động trao đổi chất của nó đặc biệt cao trong giai đoạn đầu phát triển của tôm, đòi hỏi phải lột xác nhiều lần. Chitin có thể được sử dụng làm chất bổ sung thức ăn để cải thiện tuổi thọ của tôm bị nhiễm vi khuẩn Vibrio ly giải (Cheng và cộng sự, 2021) và protein liên kết với chitin hoạt động như một bộ điều biến hoặc thụ thể nhận dạng mẫu để tạo ra phản ứng miễn dịch kháng khuẩn ở tôm (Hakimi và cộng sự, 2017).
Việc bảo vệ tôm chống lại các tác nhân lây nhiễm bắt đầu bằng phản ứng dịch thể, dựa trên các thụ thể nhận dạng mẫu (PRR) như lectin loại C (Wang XW và cộng sự, 2020) tạo ra sự tổng hợp các tác nhân miễn dịch như peptide kháng khuẩn (AMP) bằng cách nhận biết các tín hiệu kích hoạt các thụ thể giống Toll (TLR) và con đường gây suy giảm miễn dịch (IMD) (Tassanakajon và cộng sự, 2013). Ngược lại, khả năng miễn dịch tế bào được bắt đầu bởi một loạt hệ thống kích hoạt phenoloxidase (proPO), dẫn đến quá trình thực bào, đóng gói, đông máu và tạo hắc tố của mầm bệnh xâm nhập (Cerenius và cộng sự, 2008). Trong nghiên cứu này, ấu trùng Zoea bị nhiễm Vibrio parahaemolyticus và con trưởng thành cùng biểu hiện 36 gen điều hòa tăng (Bảng 2). Trong số đó, những chất liên quan đến khả năng miễn dịch tế bào bao gồm lectin loại C, peptide kháng khuẩn giả định. Lectin loại C là một loại protein nhận dạng mẫu (PRR) lớn và đã được báo cáo là có liên quan đến khả năng miễn dịch bẩm sinh của động vật không xương sống, chẳng hạn như độ bám dính tế bào, thanh lọc vi khuẩn, thực bào, kích hoạt prophenoloxidase và đóng gói (Bi và cộng sự, 2020). Các peptide kháng khuẩn (AMP) là các phân tử tác động trực tiếp để loại bỏ và loại bỏ các bệnh nhiễm trùng gây bệnh, thường được tạo ra thông qua con đường Toll và IMD, đồng thời chúng đóng một vai trò quan trọng trong khả năng miễn dịch dịch thể của giáp xác (Tassanakajon và cộng sự, 2018). Trong khả năng miễn dịch tế bào, sự xâm nhập của mầm bệnh sẽ kích hoạt hệ thống kích hoạt prophenoloxidase, bao gồm các protease serine giống trypsin nội sinh còn được gọi là chất kích hoạt phenoloxidase (PPAF) như: chất kích hoạt phenoloxidase 3-like, chất kích hoạt phenoloxidase 1-like, chất kích hoạt prophenoloxidase 2a. Sau đó, các PPAF này chuyển đổi phenol oxydase (pro-PO) thành dạng hoạt động của phenol oxydase (PO), làm mầm bệnh làm đen (Söderhäll và cộng sự, 1994). Thuốc ức chế serine protease (Serpins) là một nhóm lớn các chất ức chế protease tham gia vào nhiều quá trình sinh học quan trọng, bao gồm đông máu, tiêu sợi huyết, chết tế bào theo chương trình, phát triển và miễn dịch bẩm sinh (Kong và cộng sự, 2013). MjSerp1 đã được chứng minh là đóng vai trò tác động trực tiếp đến việc loại bỏ vi khuẩn M. japonicus (Zhao và cộng sự, 2014). Các con đường liên quan đến miễn dịch là tương tác với thụ thể ECM, nhập bào, con đường truyền tín hiệu NF-kappa B, con đường truyền tín hiệu Toll và Imd và phagosome, được thấy nhất quán ở cả ấu trùng Zoea và con trưởng thành, ngụ ý rằng khả năng miễn dịch bẩm sinh chống lại sự xâm nhập của vi khuẩn có thể tồn tại ở zoea ấu trùng, điều này phù hợp với những phát hiện của (Angthong và cộng sự, 2021).
Đối với nhóm LC-LE, đường dẫn tín hiệu MAPK, các tầng bổ sung và đông máu cũng như đường dẫn tín hiệu mTOR nằm trong số 1209 con đường liên quan đến miễn dịch được làm giàu với DEG tăng cao (P <0,05). Một số nghiên cứu đã chứng minh rằng việc kích hoạt đường truyền tín hiệu mTOR bằng cách thêm các chất vào thức ăn có thể điều chỉnh sức khỏe đường ruột của tôm (Duan và cộng sự, 2017). Ngoài ra, đường truyền tín hiệu (mTORC1) có thể ảnh hưởng đến sự nhân lên của virus đốm trắng trên tôm (Hong và cộng sự, 2022). Tuy nhiên, có rất ít nghiên cứu được thực hiện về khía cạnh chống Vibrio của tôm và cần có nghiên cứu bổ sung để xác định liệu nó có liên quan đến khả năng miễn dịch bẩm sinh ở ấu trùng tôm hay không. Tuy nhiên, ở động vật có vú, nó rất cần thiết cho tình trạng viêm ruột và hình thành tế bào biểu mô, cũng như sự phát triển và chức năng của các tế bào miễn dịch bẩm sinh và thích nghi (Mafi và cộng sự, 2022).
Các con đường được làm giàu cho các gen điều hòa giảm duy nhất tương ứng trong LC-LE và SC-SE khác nhau đáng kể. Các gen được điều hòa giảm duy nhất trong nhóm LC-LE hầu hết được làm giàu trong các con đường trao đổi chất và các con đường liên quan đến miễn dịch như hệ thống tín hiệu Notch. Ở động vật có vú, đường truyền tín hiệu Notch là đường truyền tín hiệu được bảo tồn tiến hóa cao, điều chỉnh một số quá trình sinh học như biệt hóa tế bào, phát triển mô và phản ứng miễn dịch (Lai, 2004). Sự ức chế Notch ở tế bào T trưởng thành cản trở hoạt động kháng nấm của chúng (Neal và cộng sự, 2017). Tầm quan trọng của Notch trong khả năng miễn dịch không chỉ giới hạn ở động vật có xương sống; nó đóng một vai trò quan trọng trong hệ thống miễn dịch của tôm. L. vannamei Notch (LvNotch) điều chỉnh sự hình thành các loại oxy phản ứng (ROS) trong tế bào máu và có liên quan đến phản ứng miễn dịch. Trong nghiên cứu, ấu trùng Zoea bị nhiễm Vibrio parahaemolyticus biểu hiện sự điều hòa giảm các gen liên quan đến hệ thống tín hiệu Notch, điều này phù hợp với phát hiện (Zhao và cộng sự, 2021) rằng việc làm im lặng LvNotch trong cơ thể sống đã làm tăng biểu hiện của các gen liên quan đến quá trình con đường NF-b. Con đường lysosomal là một trong những con đường miễn dịch được làm giàu cho các gen bị điều hòa ở mức độ khác nhau trong nhóm SC-SE. Lysosome duy trì cân bằng nội môi tế bào, hoạt động như trung tâm truyền tín hiệu, tích hợp quá trình trao đổi chất với quá trình thanh lọc tế bào và tham gia vào nhiều quá trình cơ bản của tế bào như sửa chữa màng tế bào, phản ứng miễn dịch và chết tế bào (Yang và Wang, 2017). Các kết quả trên chỉ ra rằng các gen và con đường miễn dịch có thể bị điều chỉnh giảm sau khi nhiễm Vibrio trong thời kỳ ấu trùng không phù hợp với các gen và con đường miễn dịch ở người trưởng thành.
5. Kết luận
Nghiên cứu của chúng tôi mô tả những thay đổi về phiên mã do nhiễm Vibrio parahaemolyticus trong giai đoạn ấu trùng Zoea hoặc trưởng thành. Chúng tôi so sánh khả năng miễn dịch chống Vibrio giữa ấu trùng Zoea và giai đoạn trưởng thành để nghiên cứu khả năng kháng bẩm sinh đối với Vibrio ở giai đoạn ấu trùng Zoea. Bằng cách phân tích dữ liệu RNA-seq, chúng tôi đã phát hiện sự hiện diện của một số con đường truyền tín hiệu liên quan đến miễn dịch như con đường truyền tín hiệu mTOR, con đường truyền tín hiệu MAPK và con đường truyền tín hiệu Notch ở giai đoạn ấu trùng Zoea, cho thấy rằng Litopenaeus vannamei có thể có khả năng tạo ra các phản ứng miễn dịch bẩm sinh chống lại sự lây nhiễm của Vibrio trong ấu trùng Zoea. Nghiên cứu này cung cấp thông tin hữu ích để hiểu rõ hơn về cơ chế miễn dịch kháng khuẩn ở ấu trùng Zoea.
Theo Nanxiong Chen, Jiaojiao Jin, Yibin Chen, Yi Cheng Hu, Yuchun Shen, Sedong Li.
Nguồn: https://www.frontiersin.org/articles/10.3389/fmars.2023.1174176/full
Biên dịch: Nguyễn Thị Quyên – Tôm Giống Gia Hóa Bình Minh
TÔM GIỐNG GIA HÓA – CHÌA KHÓA THÀNH CÔNG
Xem thêm:
- Probiotic Trong Nuôi Trồng Thủy Sản: Tầm Quan Trọng Và Triển Vọng Tương Lai
- Ảnh Hưởng Của Chế Phẩm Sinh Học Loài Bacillus (Bacillus subtilis Và B. licheniformis) Đến Chất Lượng Nước, Phản Ứng Miễn Dịch Và Sức Đề Kháng Của Tôm Chân Trắng (Litopenaeus vannamei) Chống Lại Nhiễm Nấm Fusarium solani
- Đối Mặt Với Nhu Cầu Nuôi Hải Sản Bền Vững Ngày Càng Tăng, Oman Cho Thử Nghiệm Nguồn Nước Nuôi Tôm Trên Sa Mạc