
 English
English
Tóm tắt
Hội chứng phân trắng (WFS) ở tôm nuôi được đặc trưng bởi sự xuất hiện của tôm có ruột màu trắng bất thường kết hợp với chuỗi phân trắng nổi như thảm trên bề mặt ao. Nguyên nhân của WFS rất phức tạp, nhưng một loại được gọi là EHP-WFS có liên quan đến vi bào tử trùng Enterocytozoon hepatopenaei (EHP). Gan tụy (HP), ruột và phân của tôm EHP-WFS có một lượng lớn bào tử EHP trộn lẫn với các vi khuẩn không xác định. Trong ao nhiễm EHP-WFS, một số tôm bị nhiễm EHP có biểu hiện phân trắng (WG) và tạo ra phân trắng trong khi những con tôm bị nhiễm EHP khác cùng một ao cho thấy ruột (NG) hoàn toàn bình thường và không tạo ra phân trắng. Chúng tôi đưa ra giả thuyết rằng việc so sánh hệ vi sinh vật giữa tôm WG và NG sẽ cho thấy sự kết hợp các vi khuẩn có lợi liên quan đến EHP-WFS. Để kiểm tra điều này, chúng tôi đã chọn một ao nuôi tôm thẻ Penaeus vannamei có biểu hiện WFS nghiêm trọng và sử dụng các phân tích qua kính hiển vi và vi sinh vật để so sánh các mẫu WG và NG. Về mặt mô học, EHP được xác định trong HP và ruột của cả tôm WG và NG, nhưng số lượng EHP cao hơn và tổn thương mô EHP nghiêm trọng hơn ở tôm WG. Hơn nữa, hệ vi sinh vật đường ruột của tôm WG ít đa dạng hơn và có lượng vi khuẩn phong phú hơn từ các chi Vibrio và Propionigenium. Số lượng Propionigenium trong HP của tôm WG (9364 con/100 ng DNA) cao hơn đáng kể (P = 1,1 × 10-5) so với tôm NG (12 con/100 ng DNA). Những phát hiện này đã hỗ trợ giả thuyết bằng cách phát hiện hai chi vi khuẩn ứng cử viên cần được thử nghiệm kết hợp với EHP là nguyên nhân tiềm ẩn của việc trùng phát EHP -WFS ở tôm thẻ chân trắng.
1/ Giới thiệu
Các ao nuôi tôm có biểu hiện hội chứng phân trắng (WFS), được đặc trưng bởi tôm có ruột trắng bất thường và tạo ra các chuỗi phân trắng tập trung thành các thảm nổi trên mặt ao. Hàm lượng ruột và chuỗi phân khác nhau giữa các ao bùng phát WFS, nhưng cũng chứa thành phần vi khuẩn hỗn hợp. Hai đặc điểm này chỉ ra rằng WFS có nguyên nhân phức tạp tương tự như nguyên nhân được mô tả được trong các bệnh hội chứng khác ở tôm (Kooloth Valappil và cộng sự, 2021) và đối với các bệnh động vật và thực vật nói chung (Bass và cộng sự, 2019), trong đó sự kết hợp đặc biệt của mầm bệnh (hệ sinh vật gây bệnh) có thể gây ra một bệnh cụ thể.
Một loại WFS (ATM-WFS) được đặc trưng bởi sự biến đổi lớn và bong tróc ồ ạt của các vi nhung mao từ các tế bào biểu mô ống của gan tụy (HP) của tôm thẻ chân trắng Penaeus vannamei ở Thái Lan trong điều kiện không có vi khuẩn và / hoặc vi bào tử trùng Enterocytozoon hepatopenaei (Sriurairatana và cộng sự, 2014). Trong ATM-WFS, tập hợp vi nhung mao bị bong tróc, biến đổi trong lumens hình ống dưới dạng các vật thể vermiform được gọi là vi nhung mao tổng hợp (ATM) siêu giống với gregarines. Chúng tích tụ ở cả trung tâm HP và giữa ruột và được bài tiết dưới dạng chuỗi phân trắng nổi vì hàm lượng chất béo cao. Cơ chế gây bệnh cho sự hình thành ATM vẫn còn chưa biết rõ. Sự xuất hiện của ATM-WFS là tương đối không thường xuyên vì ATM, mặc dù thường thấy (Sanguanrut và cộng sự, 2018), thường không tích lũy đủ số lượng để gây ra ATM-WFS. Ngay cả khi chúng xảy ra, ATM-WFS không liên quan đến việc gây chết số lượng nhiều hoặc các vấn đề sản xuất nghiêm trọng khác. Theo kinh nghiệm của chúng tôi, ruột trắng cũng có thể được gây ra bởi nhiễm gregarine nặng, nhiễm trùng Vibrio và bệnh HE do ăn phải tảo lam (Anjaini và cộng sự, 2018; Somboon và cộng sự, 2012). Tuy nhiên, chúng thường không dẫn đến sự tích tụ thành thảm phân trôi nổi. Do đó, các báo cáo về WFS mà không được xác nhận ít nhất bằng kính hiển vi thì không thể được quy cho một nguyên nhân cụ thể.
Gần đây một loài mới WFS đã được báo cáo ở Penaeus vannamei từ Ấn Độ (Kumar và cộng sự, 2022). Trong loại này, ATM xảy ra cùng với EHP và ở đây nó được gọi là EH/ATM-WFS. Trong các xét nghiệm sinh học trong phòng thí nghiệm, các tác giả nhận thấy rằng việc cho ăn mô HP băm nhỏ từ tôm EHP / ATM-WFS được nuôi đến tôm khỏe mạnh dẫn đến sự xuất hiện của ATM trong HP trong vòng 3 ngày sau khi cho ăn và dương tính với EHP vào ngày thứ 7 sau khi cho ăn. Tuy nhiên, WFS đã không xuất hiện cho đến sau ngày thứ 15 và chỉ kéo dài trong 3-4 ngày. Sau đó, WFS đã được giải quyết, nhưng tôm vẫn dương tính với EHP. Mặt khác, nhóm tôm được nuôi với bào tử EHP thuần chủng bằng PCR đã dương tính với EHP sau 14 ngày và vẫn bị nhiễm bệnh nhưng không có dấu hiệu WFS cho đến khi kết thúc thử nghiệm sinh học. Số lượng vi khuẩn dạng đĩa được thực hiện trong môi trường chọn lọc bằng cách sử dụng các mẫu dịch hemolymph từ WFS và tôm không WFS từ các trang trại nuôi tôm, nhưng không có kiểm tra vi khuẩn nào được thực hiện với các mẫu HP hoặc ruột tôm. Ngoài ra, không có xét nghiệm vi khuẩn nào được thực hiện trên tôm đã thử nghiệm sinh học. Tuy nhiên, trong Hình 3 của ấn phẩm của họ, phần mô HP bán mỏng từ mẫu trang trại WFS nhuộm màu xanh toluidine cho thấy rõ sự hiện diện của vi khuẩn hình que cùng với bào tử EHP và ATM. Điều này cho thấy rằng mô HP băm nhỏ từ tôm WFS được sử dụng để nuôi tôm trong thử nghiệm sinh học cũng có thể chứa vi khuẩn cùng với bào tử EHP và ATM.
Loại WFS được kiểm tra trong nghiên cứu này (EHP-WFS) không liên quan đến ATM trong ruột hoặc HP. Thay vào đó, nó được đặc trưng bởi tôm có ruột tạo ra chuỗi phân trắng chứa các tế bào gan tụy bị bong tróc, mảnh vụn mô, tế bào vi khuẩn và một lượng lớn vi bào tử trùng Enterocytozoon hepatopenaei (EHP) (Tourtip và cộng sự, 2009). Tôm EHP-WFS có biểu hiện từ trắng sang vàng, vỏ ngoài lỏng lẻo, giảm lượng thức ăn và chậm phát triển, thay đổi kích thước lớn, giảm tăng trưởng trung bình hàng ngày, tỷ lệ chuyển đổi thức ăn tăng cao và đôi khi chết. EHP-WFS đã được báo cáo từ Việt Nam trong Penaeus monodon (Hà và cộng sự, 2010), nhưng EHP không phải lúc nào cũng đi kèm với WFS và tôm có thể phục hồi sau WFS nhưng vẫn bị nhiễm EHP nặng mà không có biểu hiện WFS (Flegel, 2012; Tangprasittipap và cộng sự, 2013). Ngoài ra, một nghiên cứu về WFS ở Indonesia (Tang và cộng sự, 2016) đã báo cáo rằng “tất cả tôm được kiểm tra đều bị nhiễm EHP kèm theo hoại tử gan tụy (SHPN).” Do đó, việc thiếu bằng chứng rõ ràng về mối quan hệ nhân quả giữa EHP và WFS đã dẫn đến suy đoán rằng các loài sinh vật nhân chuẩn khác (Dai và cộng sự, 2019) hoặc các loài vi khuẩn (Chaijarasphong và cộng sự, 2020; Hou và cộng sự, 2018; Huang và cộng sự, 2020) có thể đóng nhiều vai trò trong việc gây ra WFS.
EHP-WFS hiện đang được báo cáo từ Trung Quốc (Shen và cộng sự, 2019; Wang và cộng sự, 2020), Đông Nam Á (Caro và cộng sự, 2020; Desrina và cộng sự, 2020; Flegel, 2012; Hà và cộng sự, 2010; Sajiri và cộng sự, 2021; Tang và cộng sự, 2016) và Nam Á (Prathisha và cộng sự, 2019; Rajendran và cộng sự, 2016). Nó thường xảy ra sau 30–40 ngày nuôi. Ở Thái Lan, tỷ lệ mắc cùng lúc EHP-WFS đã tăng lên đáng kể trong những năm gần đây trên tất cả các khu vực nuôi trồng thủy sản, và nó đe dọa kinh tế sản xuất tôm ở Thái Lan do kết hợp thiệt hại từ sự tăng trưởng chậm phát triển và đôi khi là gây chết.
Để điều tra khả năng nguyên nhân của EHP-WFS liên quan đến sinh vật nhân chuẩn (EHP) và vi khuẩn (loài chưa biết), chúng tôi đã tận dụng thực tế trong các đợt bùng phát EHP-WFS, một số tôm trong ao cho thấy ruột trắng (WG) trong khi những con khác cho thấy ruột (NG) hoàn toàn bình thường. Chúng tôi đưa ra giả thuyết rằng việc so sánh hệ vi sinh vật giữa tôm WG và NG từ cùng một ao EHP-WFS sẽ cho thấy sự kết hợp có thể xảy ra của các vi sinh vật có thể liên quan đến EHP-WFS. Để kiểm tra giả thuyết này, chúng tôi đã sử dụng kết hợp phân tích mô bệnh học và phân tích trình tự chuỗi amplicon 16S rRNA thông lượng cao của các hệ vi sinh vật vi khuẩn để so sánh HP và ruột của tôm WG và NG. Các phân tích so sánh của chúng tôi cho thấy các đặc điểm khác biệt giữa tôm WG và NG và cho thấy mối liên hệ đáng kể giữa EHP-WFS và các đơn vị phân loại vi khuẩn chiếm ưu thế của các chi Vibrio và Propionigenium.
2/ Vật liệu và phương pháp
2.1 Các mẫu tôm
Một ao nuôi tôm thẻ chân trắng P. vannamei có bùng phát WFS đã được chọn vì số lượng bào tử EHP cao kèm theo một số tôm chết. Ao nằm ở tỉnh Chanthaburi, Thái Lan (xem Bảng bổ sung S1) và được lót bạt PE hoàn toàn. Sau 27 ngày nuôi, tôm có trọng lượng trung bình là 8,70 g và tăng trưởng trung bình hàng ngày là 0,32 g/ngày. Các mẫu được thu thập vào ngày thứ 5 sau khi dịch WFS bùng phát. Tôm có ruột trắng (WG) và tôm có ruột (NG) hoàn toàn bình thường được lựa chọn ngẫu nhiên và được kiểm tra bằng phân tích mô bệnh học, phân tử và so sánh hệ vi sinh vật. Tổng cộng có 15 con tôm WG và 15 con NG được chọn, mỗi loại 10 con để phân tích hệ vi sinh vật và phân tử; 5 con còn lại của từng loại được dùng để kiểm tra mô bệnh học. Các mẫu được thu thập theo hướng dẫn số BT 07/2561 từ Ủy ban Chăm sóc và Sử dụng Động vật Tổ chức BIOTEC (IACUC).
2.2 Phân tích hệ vi sinh vật và phân tử của mẫu DNA
Mỗi con tôm được thu thập HP và ruột riêng biệt trong các ống 1,5 ml chứa 500 μL dung dịch đệm ly giải (50 mM Tris pH 9, 0,1 M EDTA pH 8, 50 mM NaCl, 2% SDS, 100 μg / ml proteinase K) để tách chiết DNA bằng kit tách chiết mini DNA QIAamp® (Qiagen). Việc phân loại vi khuẩn bằng giải trình tự amplicon 16S rRNA thông lượng cao được thực hiện trên các mẫu DNA của ruột trên từng con tôm riêng lẻ, trong khi các phản ứng chuỗi polymerase định lượng (qPCR) được thực hiện với các mẫu DNA của HP và ruột riêng biệt từ mỗi mẫu tôm (xem Bảng bổ sung S2).
2.3 Phân tích mô bệnh học
Các mẫu tôm được chuẩn bị để kiểm tra mô học bằng các phương pháp tiêu chuẩn (Bell và Lightner, 1988). Chúng được cố định một thời gian ngắn trong dung dịch cố định AFA của Davidson trong 18–24 giờ trước khi chuyển sang ethanol 70% trước khi xử lý mô, nhúng vào parafin, cắt lớp (dày 4 μm) và nhuộm bằng hematoxylin và eosin (H&E). Các lớp đã được kiểm tra bằng cách sử dụng kính hiển vi Leica DM 750 được trang bị máy ảnh kỹ thuật số Leica ICC50 W.
2.4 Giải trình tự amplicon 16S rRNA thông lượng cao
Các mẫu DNA đã được gửi để kiểm tra chất lượng, chuẩn bị ở thư viện Illumina và giải trình tự tại Macrogen, Inc. (Hàn Quốc). Các amplicon được phân lập từ vùng biến đổi V3-V4 của vi khuẩn 16S rRNA bằng cách sử dụng mồi thuận (5′-CCTACGGGNGGCWGCAG-3′) và ngược lại (5′-GACTACHVGGGTATCTAATCC-3′) (Herlemann và cộng sự, 2011) và được sử dụng để chuẩn bị thư viện theo trình tự với kit Herculase II Fusion DNA Polymerase Nextera XT V2. Nồng độ và phân bố kích thước nguồn dữ liệu đã được định lượng trên TapeStation D1000 và sau đó được giải trình tự trên nền tảng Illumina MiSeq bằng cách sử dụng định dạng đầu cuối ghép nối 2×300. Các lần đọc trình tự đã được gửi cùng với số gia nhập NCBI BioProject PRJNA735923 trong cơ sở dữ liệu NCBI BioProject.
2.5 Phân tích quần xã vi sinh vật
Các lần đọc trình tự thô đã được cắt bớt để loại bỏ các chuỗi mồi bởi Cutadapt (https://cutadapt.readthedocs.io/) và sau đó được xử lý bằng QIIME2 (phiên bản 2019.7.0) (Bolyen và cộng sự, 2019) với dada2 denoise-paired (Callahan và cộng sự, 2016) với độ dài cắt ngắn lần lượt là 280 và 235 cặp cơ sở để đọc xuôi và ngược để tạo ra một tập hợp các biến thể chuỗi amplicon (ASV). Phân loại ASV được thực hiện bằng USearch dựa trên cơ sở dữ liệu RDP (Edgar, 2010). ASV đã được nhập vào R và được sử dụng cho các ASV được tìm thấy trong ≥ 2 mẫu và có ≥ 1% hoặc ≥ 0,1% để phân tích thêm. Các phân tích ASV với mức dồi dào ≥ 1% được trình bày trong văn bản, trong khi các phân tích về ASV dồi dào 0,1% ≥ được đưa ra trong Thông tin bổ sung. Các bộ ASV đã lọc được xử lý bằng phương pháp phân tích dữ liệu tổng hợp (CoDa) (Gloor và cộng sự, 2017) để kiểm tra tỷ lệ giữa các ASV hoặc phân tích dữ liệu đếm tiêu chuẩn. Đối với phương pháp CoDa, các ASV không đếm được đã được thay thế bằng gói zCompositions R (Palarea-Albaladejo và Martín-Fernandez, ‘ 2015), được chuyển đổi với phép biến đổi tỷ lệ logarit trung tâm sau đó một phân tích giá trị đơn lẻ (SVD) đã được áp dụng cho các biểu đồ phân tích thành phần chính (PCA) với khoảng cách của Aitchison. Các bài kiểm tra độ đa dạng khác biệt cho ASV được thực hiện với chất dẫn sinh học ALDEx2 v1.6.0 bằng cách sử dụng ASV đáng kể với sự khác biệt về kích thước hiệu ứng dự kiến là ≥ 1 (Fernandes và cộng sự, 2014). Để phân tích dữ liệu đếm tiêu chuẩn, chúng tôi đã sử dụng các gói phyloseq (McMurdie và Holmes, 2013) và microbiome (https://microbiome.github.com/microbiome) để tính toán chỉ số đa dạng alpha và tỷ lệ đa chiều phi hệ mét (NMDS) với khoảng cách không giống nhau Bray-Curtis và DESeq2 (Love và cộng sự, 2014) cho phép thử mức độ chênh lệch (FDR < 0,05). Phân tích đa biến hoán vị của phương sai (adonis và anosim; 10.000 hoán vị), tính đồng nhất của phân tán đa biến (betadisper với permutest 10.000 hoán vị và thử nghiệm của Tukey) và đồ họa bổ sung đã được thực hiện với các gói Vegan (https://github.com/vegandevs/thuầnchay), hệ thống ggplot2 (https://ggplot2.tidyverse.org ) và gói ggpubr (https://rpkgs.datanovia.com/ggpubr/)
2.6 Định lượng phân tử bằng các phản ứng chuỗi polymerase
PCR định lượng (qPCR) đã được sử dụng để định lượng số lượng vi bào tử trùng của EHP và chọn các đơn vị phân loại Propionigenium trên 100 ng tổng số DNA được chiết xuất. Như đã thảo luận ở phần sau bản thảo này, không có mồi PCR phù hợp nào có thể được thiết kế để xác định các loài Vibrio cụ thể dựa trên trình tự ASV biểu hiện quá mức trong các mẫu WG. Mỗi phản ứng qPCR được thực hiện với tổng thể tích 20 μL, bao gồm 10 μL 2X KAPA SYBR FAST qPCR Master Mix (KAPA Biosystems, Hoa Kỳ), 0,2 μM mồi thuận, 0,4 μL Low ROX, 100 ng mẫu DNA và nước cất đến tổng thể tích 20 μL. Mồi cho EHP đã được mô tả trong Jaroenlak và cộng sự (2016) và trong Kanitchinda và cộng sự (2020), trong khi những đơn vị phân loại Propionigenium được thiết kế dựa trên kết quả giải trình tự của nghiên cứu này – PG16S-F (5′-TGGACAATGGACCAAAAGTCTG-3′) và PG16S-R (5′- TGTAGTTCCCGCCCTCTCTG – 3′). Các mẫu DNA để tạo đường cong tiêu chuẩn được lấy từ các đoạn mục tiêu đã được tinh sạch có cùng một bộ mồi tương ứng với số lượng vi bào tử trùng ước tính khác nhau của dung dịch pha loãng gấp mười lần từ 108 đến 102 bào tử / 1 μL. Khuếch đại cho phép đo qPCR được thực hiện bằng Hệ thống PCR thời gian thực nhanh 7500 (Hệ thống sinh học ứng dụng, Hoa Kỳ) với các điều kiện sau: đối với EHP, 2 phút ở 94ºC, tiếp theo là 40 chu kỳ 30 giây ở 94ºC, 30 giây ở 64ºC và 30 giây ở 72ºC; và đối với đơn vị phân loại Propionigenium, 3 phút ở 95ºC, tiếp theo là 40 chu kỳ 15 giây ở 95ºC, 30 giây ở 55ºC, và 30 giây ở 72ºC. Kiểm soát không theo mẫu và DNA của HP, ruột và các đường cong tiêu chuẩn đã thu được trong các phản ứng ba lần. Các phân tích đường cong nóng chảy và đường cong tiêu chuẩn đã đánh giá tính đặc hiệu của các phản ứng để thu được số lượng bào tử ước tính với đường cơ sở được chỉ định phần mềm tự động và ngưỡng được đặt thủ công ở mức 0,145 bằng cách sử dụng phần mềm Hệ thống phát hiện trình tự ABI PRISM® 7500 (v2.3). Thử nghiệm Mann–Whitney U được thực hiện theo R và sự khác biệt ở p ≤ 0,05 được coi là dấu hiệu thống kê (xem Bảng bổ sung S3).
3/ Kết quả
3.1 Các dấu hiệu lâm sàng trong ao WFS và mô bệnh học của các đường tiêu hóa của tôm
Tôm WG có đường tiêu hóa trong đó dạ dày, HP và toàn bộ ruột tôm có màu trắng. Chúng bị mềm vỏ hoặc vỏ lỏng lẻo. Ngược lại, tôm NG hoàn toàn bình thường (Hình bổ sung. S1). Kiểm tra mô bệnh học của tôm WG và NG cho thấy cả những bất thường chung và riêng biệt. Các bất thường gặp bao gồm các tế bào bị teo và bào tử EHP trong các tế bào biểu mô ống HP và trong các tế bào biểu mô của phần ruột được bao quanh bởi bào tử của HP (Hình 1 và 2). Các tổn thương khu trú bao gồm tế bào tạo máu của tôm bao bọc các bào tử EHP tổng hợp cũng được quan sát thấy ở cả hai nhóm (Hình 1A). Các đặc điểm cụ thể được quan sát thấy ở tôm WG là 1) lượng bào tử EHP cao hơn trong các tế bào biểu mô HP và ruột (Hình 1 và 2), và 2) lượng bào tử EHP tự do cao hơn, tế bào HP bị bong tróc và tế bào vi khuẩn hình que trong lòng HP (Hình 2A và phần trong). ATM đôi khi được tìm thấy ở tôm bình thường và trong tôm WFS-ATM (Sriurairatana và cộng sự, 2014) không có trong HP hoặc ruột của tôm được kiểm tra trong nghiên cứu này (xem Hình 1 và 2).
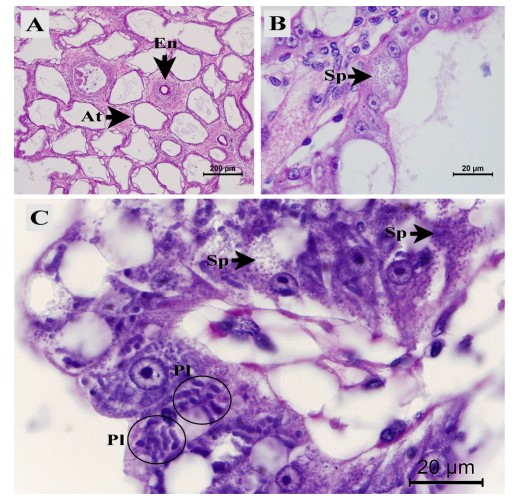
Hình 1. ảnh chụp vi thể của các đặc điểm mô học của các mô gan tụy ruột trắng (WG) và ruột (NG) bình thường. Đặc điểm tương tự của tôm WG và NG là (A) tế bào bị teo (At) của ống biểu mô gan tụy và bao bọc tế bào máu (En), (B) bào tử EHP (Sp) trong tế bào biểu mô gan tụy và (C) tỷ lệ hiện mắc plasmodia (Pl) và bào tử (Sp) cao trong biểu mô ống gan tụy.
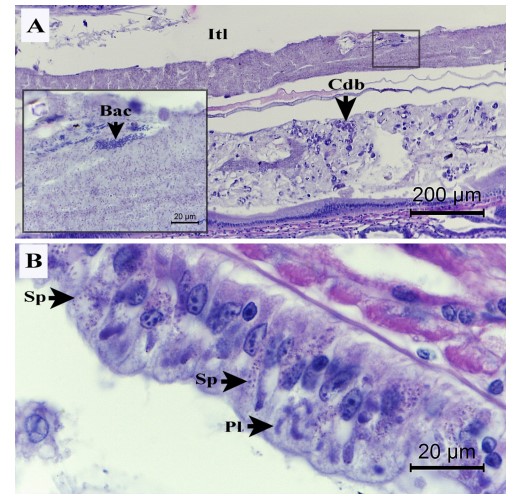
Hình 2. Ảnh chụp vi thể của ruột tôm phân trắng. (A và inset) Lumen (Itl) chứa các mảnh vụn tế bào biểu mô HP (Cdb), khuẩn lạc của vi khuẩn hình que (Bac) và khối bào tử Enterocytozoon hepatopenaei (EHP). (B) Bào tử (Sp) và plasmodia (Pl) của các tế bào biểu mô bị nhiễm EHP ở ruột. Lưu ý rằng biểu mô giữa tương đối bình thường và nguyên vẹn, mặc dù có sự hiện diện của các giai đoạn EHP trong một số tế bào.
3.2 So sánh hệ vi sinh vật đường ruột giữa tôm WG và NG
Các cặp đọc thô (5,974,443) của các mẫu DNA ruột riêng lẻ đã tạo ra 2,254 biến thể trình tự amplicon (ASV) với khử nhiễu QIIME2 DADA2. Các ASV này được phát hiện trong ≥ 2 mẫu và sự phong phú ≥ 1% hoặc ≥ 0,1%. Kiểm tra ban đầu về hai bộ dữ liệu được phân tách bằng phân tích thành phần chính (PCA) và tỷ lệ đa chiều phi hệ mét (NMDS) cho thấy các mẫu WG và NG có các loại vi khuẩn khác nhau, ngoại trừ một mẫu WG (F8) được nhóm lại chặt chẽ với các mẫu NG (Hình bổ sung. S2, S3, S4 và S5; và Vật liệu và Phương pháp). PCA dựa trên dữ liệu biến đổi logarit tập trung của các mẫu và tải trọng liên quan cho ≥ 1% dữ liệu ASV phong phú (Hình 3) cho thấy các cộng đồng vi khuẩn đường ruột giữa tôm WG và NG khác nhau rõ rệt, ngoại trừ một mẫu tôm WG (F8) tương tự như nhóm NG hơn. Sự khác biệt của các cộng đồng vi khuẩn đường ruột giữa tôm WG và NG đã được cho là dấu hiệu thống kê với khoảng cách Bray-Curtis (adonis, phương sai 32,5%, P = 9,9 × 10-5) và Aitchison (anosim, R = 0,86, Signifcant = 1 × 10-3). Chúng tôi đã bao gồm tất cả các mẫu WG và NG vào các phân tích tiếp theo của mình, mặc dù mức độ nghiêm trọng cao hơn đã được quan sát thấy khi loại trừ mẫu WG F8 (dữ liệu không được hiển thị).
Độ đa dạng alpha trong WG thấp hơn đáng kể so với các mẫu NG (chỉ số Độ Richness của Chao1, P = 2 × 10-4; Chỉ số đa dạng của Shannon, P = 1, 4 × 10-4; Chỉ số Gini-Simpson, P = 5, 8 × 10-2; Hình 4). Trung bình, khoảng cách giữa các mẫu WG (trên PCA và NMDS) tương đối lớn hơn so với các mẫu NG (betadisper với khoảng cách BrayCurtis, P = 5,8 × 10-2 từ cả thử nghiệm permutest và Tukey và với khoảng cách của Aitchison, lần lượt là P = 2,5 × 10-2 và 2,7 × 10-2 từ các thử nghiệm permutest và Tukey; Hình 3 và bổ sung. S2, S3, S4 và S5), cho thấy sự khác biệt nhiều hơn trong các quần thể vi khuẩn giữa các loài tôm riêng lẻ trong nhóm WG so với các loài trong nhóm NG, tức là nhóm WG không đồng nhất hơn.
Để xác định các đơn vị phân loại vi khuẩn liên quan đến tôm EHP-WFS, chúng tôi đã phân tích các ASV biểu hiện quá mức trong các mẫu WG (xem Vật liệu và Phương pháp). ASV của các chi Vibrio và Propionigenium trên các mẫu tôm WG khác nhau đáng kể so với các mẫu NG (Hình 3 và Hình 5; xem Vật liệu và Phương pháp). Những thay đổi trung bình về độ siêu đa dạng của các mẫu WG so với các mẫu NG cho các ASV chi Vibrio lần lượt là 34,3–51,3 (3 ASV) và 6,6 × 107 (1 ASV) từ Aldex2 và DESeq2, và sự thay đổi đối với chi Propionigenium lần lượt là 134 – 934 (2 ASV) và 1,7 × 107 (1 ASV) từ Aldex2 và DESeq2 (Hình 5). Lưu ý rằng tất cả các ASV thuộc chi Propionigenium trong bộ dữ liệu hệ vi sinh vật đường ruột đều là ASV được mô tả quá mức trong các mẫu WG. Các ASV tương tự liên quan đến các đơn vị phân loại Vibrio và Propionigenium này và một số chi bổ sung cũng được thu thập cùng với các bộ dữ liệu ASV khác (xem Vật liệu và Phương pháp và Bảng bổ sung S4).
Chúng tôi tập trung vào các ASV đại diện đáng kể WG của Vibrio và Propionigenium để điều tra thêm về tầm quan trọng của chúng trong EHP-WFS. Trình tự của các ASV Propionigenium đại diện quá mức trong các mẫu WG đều khớp với P. maris có độ nhận dạng cao (99,75%; Số GenBank Accession NR_026243.1 và Y16800.1) hoặc giống với số vi khuẩn chưa được nuôi cấy được xác định là Propionigenium sp., nhưng có khả năng là phân lập của P. maris. Do đó, các chuỗi mồi cụ thể được sử dụng có thể được thiết kế để so sánh sự phong phú của Propionigenium trong ruột tôm và HP. Tuy nhiên, các chuỗi Vibrio ASV được đại diện nhiều WG đều phù hợp với các bản ghi của nhiều loại của nhánh Vibrio harveyi do vùng rRNA 16S ngắn được nhắm mục tiêu. Các ASV Vibrio không được đánh giá quá cao, của WG cũng vậy. Do đó, không thể tạo ra một cặp mồi đặc trưng loài để định lượng so sánh các loài Vibrio được đại diện WG. Điều này không có nghĩa là chúng tôi coi sự tham gia Vibrio là một trong những nguyên nhân không đáng kể của WFS, chỉ là chúng tôi không thể, từ dữ liệu của mình, xác định một hoặc nhiều loài cụ thể để phân tích thêm.
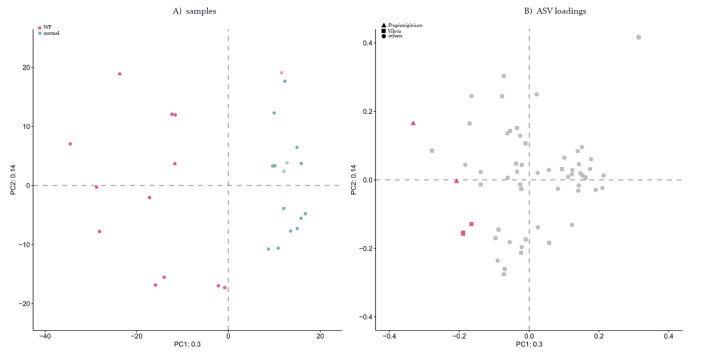
Hình 3. Biểu đồ PCA thành phần của các mẫu (A) và tải ASV (B) cho bộ dữ liệu ASV dồi dào 1% ≥ (Vật liệu và Phương pháp). Trong bảng A, mỗi điểm là một mẫu [tô màu cho nhóm tôm WG (đỏ) và NG (xanh); Bảng S1] và khoảng cách giữa các điểm tỷ lệ thuận với chênh lệch đa biến giữa các mẫu. Bảng B hiển thị tải cho bảng A trong cùng một không gian tọa độ, thể hiện sự đóng góp của ASV vào việc tách các mẫu. Trong biểu đồ này, mỗi điểm là một ASV (được định hình bởi chi phân loại và được tô màu bởi các ASV phong phú cao hơn đáng kể được chỉ định trong nhóm WG (màu đỏ) và khoảng cách và hướng từ nguồn gốc đến điểm đại diện cho ASV tỷ lệ thuận với độ lệch chuẩn của ASV đó trong tập dữ liệu. Khoảng cách giữa ASV này và ASV khác tỷ lệ nghịch với liên kết thành phần của chúng: các điểm gần nhau có thể có sự phong phú tương đối phù hợp trên tất cả các mẫu. Khả năng diễn giải trực tiếp cốt truyện bị giới hạn bởi tỷ lệ phương sai được giải thích (30% trên thành phần đầu tiên và 14% trên thành phần thứ hai).
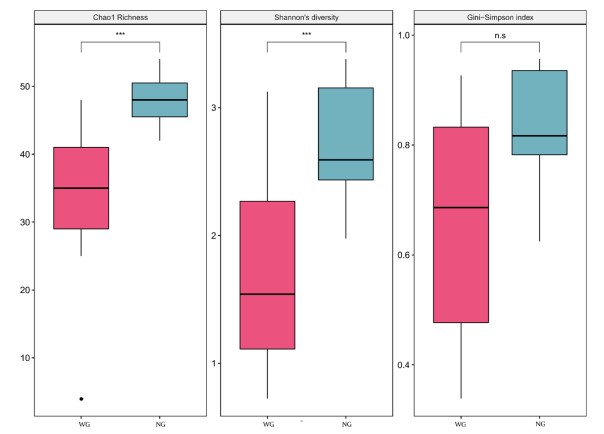
Hình 4. So sánh sự đa dạng alpha giữa nhóm tôm WG (n = 13, màu đỏ) và nhóm tôm NG (n = 15, màu xanh lá cây) với chỉ số Độ phong phú của Chao1, chỉ số đa dạng của Shannon và chỉ số Gini-Simpson. Sự khác biệt đáng kể được đưa ra bởi dấu hoa thị (n.s., P ≥ 0.05; ***, 0.0001 ≤ P < 0.001).
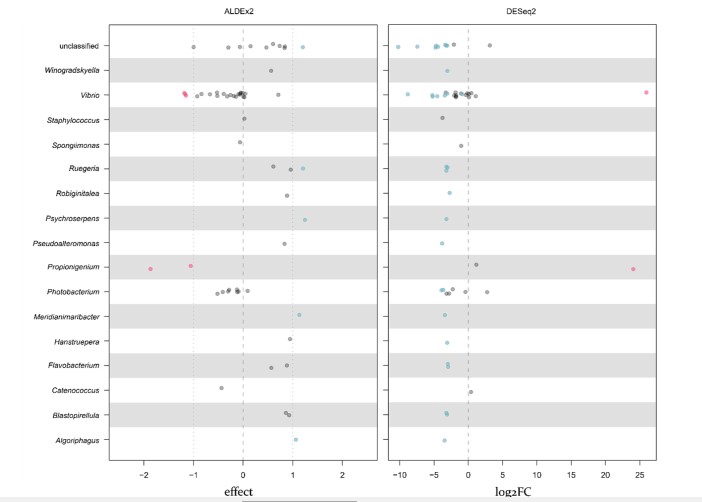
Hình 5. Sự phong phú tương đối khác biệt của ASV được ngăn bởi chi được xác định bởi (A) ALDEx2 và (B) DESeq2. Các điểm có màu đỏ hoặc xanh lam nếu chúng có nhiều đặc điểm trong nhóm tôm WG hoặc NG, tương ứng (Vật liệu và Phương pháp).
3.3 Mức độ propionigenium cao trong gan tụy của nhóm tôm WG nhưng không có trong nhóm NG
Để so sánh mật độ Propionigenium và EHP giữa tôm WG và NG ở cả HP và ruột, qPCR được thực hiện trên các mẫu DNA HP và ruột riêng lẻ cho từng mẫu tôm. Số lượng bào tử trên 100 ng DNA cho cả EHP và Propionigenium trong tôm WG và NG trong HP và ruột được thể hiện trong Hình 6 và Bảng bổ sung S2. Mặc dù có hệ số biến động cao (CV = 0,9 đến 1,4) của số lượng bào tử đối với cả Propionigenium và EHP trong cả bốn nhóm được sử dụng để so sánh, một số khác biệt có dấu hiệu thống kê đã được nhìn thấy (Bảng bổ sung S3).
Trong HP, số lượng bào tử Propionigenium trong tôm WG cao hơn đáng kể so với tôm NG (phương tiện lần lượt là 8.487/100 ng so với 9/100 ng DNA đối với WG và NG; P = 2,2 × 10-5, thử nghiệm Mann–Whitney U; Hình 6A). Phạm vi định lượng cho tất cả HP của các mẫu tôm NG là từ DNA không thể phát hiện (0/100 ng) đến 41/100 ng DNA. Những con số thấp này được ước tính bằng cách mở rộng vượt quá điểm nồng độ thấp nhất là 100/100 ng trong đường cong tiêu chuẩn. Ngược lại, phạm vi của nhóm WG là 109 đến 22.837/100 ng DNA. Những kết quả này chỉ ra rằng nhóm WG mang tải lượng Propionigenium trong HP cao hơn đáng kể so với nhóm NG.
Đối với EHP trong HP, nhóm tôm WG có số lượng bào tử trung bình trên 100 ng DNA cao hơn đáng kể so với tôm NG (lần lượt là 61.775 so với 34.333/100 ng DNA; P = 0,04, thử nghiệm Mann–Whitney U; Hình 6A). Phạm vi số lượng trên 100 ng DNA trong nhóm WG là 2.711–184.413 (CV = 0,9) trong khi đối với nhóm NG là 790–162.461 (CV = 1,3). Những kết quả này chỉ ra rằng tôm WG có xu hướng có tải lượng EHP trong HP cao hơn tôm NG, cho thấy bào tử EHP cao trong HP có thể liên quan đến WFS.
Trong các mẫu ruột, Propionigenium có số lượng bào tử trung bình cao hơn đáng kể trên 100 ng DNA trong nhóm WG so với nhóm NG (có nghĩa là 21.035 so với 859/100 ng DNA; P = 0,05, thử nghiệm Mann–Whitney U; Hình 6B). Phạm vi số lượng trên 100 ng DNA cho nhóm WG là 1 đến 79.650 (CV = 1,4), trong khi ở nhóm NG là 103–3.345 (CV = 1,2). Một lần nữa, kết quả cho thấy tôm WG có xu hướng chứa nhiều Propionigenium ở ruột hơn tôm NG.
Đối với EHP ở ruột, không có sự khác biệt rõ rệt về số lượng giữa tôm WG và NG (lần lượt là 15.940 và 16.322 /100 ng DNA; P = 0,44, thử nghiệm Mann–Whitney U; Hình 6B). Phạm vi số lượng trên 100 ng DNA cho nhóm WG là 370 đến 60.597 (CV = 1,0), trong khi ở nhóm NG là 415 đến 83.259 (CV = 1,5).
Từ các xét nghiệm qPCR của chúng tôi, số bào tử Propionigenium cũng được lập biểu đồ dựa trên số bào tử EHP từ các HP của mẫu tôm (Hình 6C). Điều này cho thấy sự xuất hiện đồng thời của Propionigenium với các bào tử EHP cao trong tôm WG và sự xuất hiện đồng thời của Propionigenium và các bào tử EHP thấp hơn trong tôm NG (Hình 6A và 6C).
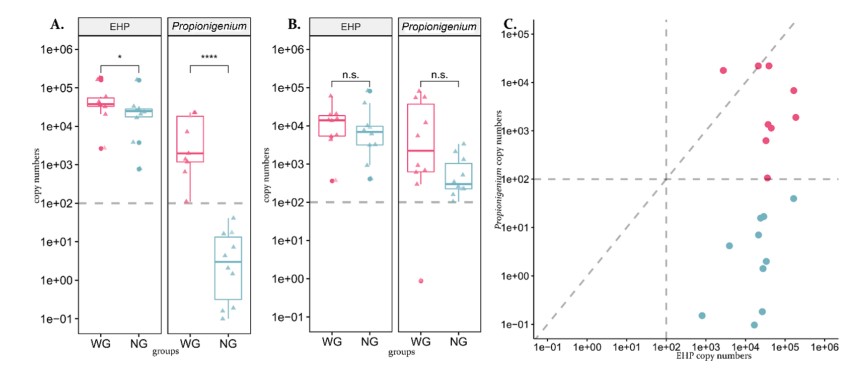
Hình 6. Sao chép số lượng Propionigenium và EHP trong 100 ng mẫu DNA của heapatopancreas (A) và midguts (B) giữa các nhóm WG và NG, và phân tán (C) của số bản sao của Propionigenium so với số của EHP trong các mẫu gan tụy. Các điểm được tô màu đỏ và xanh lam cho các nhóm WG và NG, tương ứng. Số lượng bản sao ước tính thu được bằng các phản ứng qPCR được mô tả trong văn bản.
4/ Thảo luận
Chúng tôi chỉ ra rằng sự xuất hiện đồng thời của EHP và các cộng đồng vi khuẩn cụ thể có thể góp phần như một bệnh lý sinh vật nhân sơ-nhân chuẩn có thể gây ra biểu hiện lâm sàng của WFS ở tôm he. Tôm có các tập đoàn vi sinh vật xuất hiện đồng thời này trong HP thể hiện đặc tính ruột trắng (WG) của WFS, trong khi những con khác được thu thập từ cùng một ao nhưng có màu ruột bình thường (NG) thì không có bệnh lý này ở HP.
Đối với mô bệnh học của ruột, chúng tôi đã kết hợp các báo cáo trước đó rằng tôm WG có tổn thương HP nghiêm trọng hơn, đặc trưng bởi số lượng bào tử cao hơn và phá hủy mô nhiều hơn (ví dụ: tế bào bị tách, teo và bong tróc) so với tôm NG (Hình 1 và 2). Ngoài ra, số lượng lớn các tế bào biểu mô có chứa plasmodia và/hoặc bào tử EHP chỉ được quan sát thấy ở tôm WG trong vùng ruột đi qua HP. Các cuộc kiểm tra của chúng tôi cũng đã kết hợp các báo cáo trước đó về các tế bào vi khuẩn hình que mang các bào tử EHP (Tang và cộng sự, 2016; Tangprasittipap và cộng sự, 2013, Thitamadee và cộng sự, 2016).
Trong một số mẫu WG và NG, các mô HP cho thấy các tổn thương với sự kết tụ và bao bọc tế bào máu (Hình 1A). Từ những cảm nhiễm trong phòng thí nghiệm với EHP, người ta đã chứng minh rằng không có phản ứng tan máu với EHP đơn lẻ (Salachan và cộng sự 2017; Chaijarasphong và cộng sự, 2020), vì vậy chúng tôi giả định rằng sự kết tụ hồng cầu và u hạt kèm theo sự hiện diện của vi khuẩn trong HP không phải do EHP gây ra. Thật vậy, ký sinh trùng nội bào thường không tiết ra huyết cầu. Ví dụ, vi khuẩn Agmasoma penaei trong mô cơ P. monodon hiếm khi xảy ra (Flegel và cộng sự, 1992). Có lẽ một phản ứng viêm có thể được kích hoạt do tổn thương mô hoặc ly giải tế bào, dẫn đến giải phóng các kháng nguyên ký sinh trùng nội bào. Trong nhóm ký sinh trùng, một số loài côn trùng như bộ cánh vẩy lepidoptera và bộ cánh thẳng orthoptera có dấu hiệu miễn dịch tế bào do tăng số lượng tế bào máu, thực bào, bao bọc, hình thành nốt sần và melanize trong các mô bị nhiễm bệnh (Hoch và cộng sự, 2004; IaL và cộng sự, 2004; IuIa và cộng sự, 2000; Tokarev và cộng sự, 2007). Tuy nhiên, một số loài microsporidia có thể thoát khỏi hoặc ngăn chặn khả năng miễn dịch của vật chủ theo lợi thế của chúng (Antúnez và cộng sự, 2009). Kết quả kiểm tra mô bệnh học của chúng tôi cũng cho thấy sự tích tụ tế bào vi khuẩn hình que cao hơn trong lòng ruột ở WG so với tôm NG, cho thấy sự liên quan của vi khuẩn kết hợp với EHP trong việc gây ra EHP-WFS
Chúng tôi cũng nhận thấy rằng HP của tôm WG có tải lượng EHP cao hơn đáng kể so với tôm NG, được đo bằng xét nghiệm qPCR (lần lượt là 61.775 so với 34.333/100 ng DNA) cho thấy tải lượng EHP cao có liên quan đến WFS. Tuy nhiên, một số tôm NG có số lượng (ví dụ: 1,6 × 105/100 ng DNA) gần bằng số lượng cao nhất trong nhóm WFS (1,8 × 105), cho thấy rằng chỉ riêng vi khuẩn EHP cao không đủ để gây ra WFS. Điều này phù hợp với các nghiên cứu chung sống EHP của chúng tôi và với việc cho ăn trực tiếp các bào tử EHP trong phòng thí nghiệm, nơi các bào tử EHP cao từ 1,1 × 105 đến 3,6 × 106 trên 100 ng DNA đã được ghi nhận (Munkongwongsiri và cộng sự, 2021) nhưng chưa bao giờ được liên kết với WFS. Do đó, mối liên quan với WFS vẫn chưa rõ ràng và có thể, tải lượng và bệnh lý EHP cộng với một hoặc nhiều yếu tố môi trường có thể liên quan đến biểu hiện của nó.
Nghiên cứu gần đây sử dụng lysozyme lòng trắng trứng gà mái (HEWL) cho thấy rằng khẩu phần ăn bổ sung HEWL có thể ngăn ngừa WFS và cải thiện tỷ lệ sống và tăng trưởng của tôm trong ao nuôi. HEWL có hoạt tính kháng khuẩn chống lại một số loài Vibrio và cho ăn bằng khẩu phần ăn bổ sung HEWL dẫn đến việc loại bỏ các tế bào Vibrio trong đường tiêu hóa của tôm (Woraprayote và cộng sự, 2020). Phân tích chuỗi amplicon 16S rRNA thông lượng cao của chúng tôi cho thấy vi khuẩn thuộc chi Vibrio và Propionigenium có liên quan đáng kể với tôm WG (Hình 3 và 5). Sau đó, theo qPCR cho rằng nồng độ Propionigenium trong các mẫu HP và ruột của WG cao hơn nồng độ trong tôm NG (Hình 6 và Bảng bổ sung S2). Những so sánh tương tự không thể được thực hiện với các loài Vibrio ưu thế, các chuỗi trong số đó đều liên quan đến chi Vibrio harveyi (Darshanee Ruwandeepika và cộng sự, 2012; Ke và cộng sự, 2017; Urbanczyk và cộng sự, 2013). Điều này là do các mồi được sử dụng để khuếch đại chung của 16S rRNA tạo ra amplicon quá ngắn và quá giống nhau để phân biệt các loài Vibrio riêng lẻ trong một nhánh. Về mặt này, chúng tôi không hạ thấp vai trò của đơn vị phân loại Vibrio trong bệnh lý của EHP-WFS lâm sàng và vấn đề này nên là chủ đề của nghiên cứu sâu hơn. Những mối liên hệ giữa vi khuẩn của chi Propionigenium và Vibrio với EHP-WFS đã được quan sát và hỗ trợ bởi cả phân tích amplicon 16S rRNA thông lượng cao và phân tích qPCR.
Sự phong phú ngày càng tăng của Vibrio spp. cơ hội (được kiểm tra bằng phương pháp phết đĩa truyền thống) đã được báo cáo trong các ao WFS của cả tôm sú P. monodon và P. vannamei ở nhiều nước châu Á. Cụ thể, các chủng Vibrio được báo cáo từ ruột tôm WFS và nước nuôi là V. harveyi, V. alginolyticus, V. parahaeolyticus, V. anguillarum, V. fluvialis, V. mimicus, V. vulnifcus, V. damselae và V. cholera (Aranguren Caro và cộng sự, 2021; Huang và cộng sự, 2020; Somboon và cộng sự, 2012; Supono và cộng sự, 2019; Wang và cộng sự, 2020). Một số chủng của V. sinaloensis và V. parahaemolyticus đã chứng minh độc lực thông qua khả năng gây chết tôm trong các xét nghiệm sinh học thử nghiệm gần đây, nhưng không có dấu hiệu lâm sàng WFS (Wang và cộng sự, 2020). Nhưng gần đây, một chủng cụ thể của V. parahaemolyticus có thể tạo ra các dấu hiệu lâm sàng WFS bằng thực nghiệm cảm nhiễm tôm bị nhiễm EHP nặng (Aranguren Caro và cộng sự, 2021). Thí nghiệm đồng nhiễm V. parahaemolyticus và EHP hỗ trợ bệnh lý sinh vật nhân sơ-nhân chuẩn như các tác nhân gây bệnh kết hợp cho biểu hiện lâm sàng của EHP-WFS. Sử dụng các phương pháp không phụ thuộc vào nuôi cấy để giải trình tự amplicon thông lượng cao hoặc giải trình tự hệ thống siêu âm bằng shotgun, các nghiên cứu WFS gần đây đã kiểm tra xem các tổ hợp cộng đồng vi sinh vật đường ruột WFS có khác với tôm khỏe mạnh hay không. Trong các ao P. Vannamei nhiễm WFS ở Trung Quốc và Indonesia, các nghiên cứu gần đây về hệ vi sinh vật WFS cho thấy cấu trúc khác biệt rõ rệt của hệ vi sinh vật đường ruột WFS chuyển sang “rối loạn sinh lý” đường ruột ít sự đa dạng hơn so với tôm khỏe mạnh nhưng không đồng nhất thành phần vi khuẩn (Alfansah và cộng sự, 2020; Hou và cộng sự, 2018; Huang và cộng sự, 2020; Wang và cộng sự, 2020). Rối loạn sinh lý đường ruột tôm đã được quan sát thấy trong một số nghiên cứu EHP-WFS (Wang và cộng sự, 2020), nhưng các nghiên cứu khác thì không điều tra sự tồn tại này (Alfansah và cộng sự, 2020; Hou và cộng sự, 2018; Huang và cộng sự, 2020) . Quan trọng là, các ứng cử viên vi khuẩn quan trọng liên quan đến WFS thu được bằng các phân tích thống kê cho thấy các đơn vị phân loại vi khuẩn dồi dào hơn đáng kể trong WFS so với tôm bình thường. Chúng bao gồm các đơn vị phân loại có liên quan đến Vibrio, Candidatus Bacilloplasma, Aeromonas, Phascolarctobacterium, Ruminococcus, Rhodobacteraceae, Alteromonas, Marinomonas, Photobacterium, Pseudoalteromonas và Flavobacteraceae (Alfansah và cộng sự, 2020; Hou và cộng sự, 2018; Huang và cộng sự, 2020; Wang và cộng sự, 2020). Các phân tích hệ vi sinh vật của chúng tôi (Hình 3 và 4) đã hỗ trợ các đặc tính của sự đa dạng vi khuẩn thấp hơn trong các mẫu WG và chuyển các thành phần hệ vi sinh vật đường ruột sang rối loạn chức năng đường ruột ở tôm WG. Nghiên cứu của chúng tôi đã bổ sung một loại vi khuẩn thuộc chi Propionigenium vào danh sách các vi khuẩn liên quan đến WFS, đặc biệt là trong các ao EHP. Các ao WFS có biểu hiện tôm chết bất thường.
Điều đặc biệt quan tâm là sự khác biệt rõ rệt về sự hiện diện thấp hoặc không có Propionigenium trong HP của tôm NG nhưng sự hiện diện đáng kể trong HP của tôm WG trong nghiên cứu của chúng tôi (Hình 6A). Có thể sự tiến triển của NG thành tôm WG có thể liên quan đến sự di chuyển của Propionigenium (có lẽ cùng với Vibrio) từ ruột vào HP.
HP của tôm khỏe mạnh thường không có vi khuẩn và sự hiện diện của vi khuẩn trong HP biểu thị tình trạng sức khỏe kém (ví dụ: Vibriosis (Lightner, 1996)). Chi Propionigenium trước đây không liên quan đến bệnh tôm. Cho đến nay, nó bao gồm hai loài vi khuẩn kỵ khí nghiêm ngặt (P. maris và P. modestum) có khả năng khử cacboxyl hóa succinate thành propionate để hỗ trợ cho sự tăng trưởng (Schink, 2006). Chúng được tìm thấy trong môi trường biển, điển hình là trong lớp trầm tích. Chúng là các tế bào gram âm, cầu khuẩn hoặc giống như que ngắn với các đầu tròn (Schink, 2006). Trong số hai loài hiện đang được biết đến, trình tự amplicon 16S rRNA ngắn của chúng tôi cho thấy sự tương đồng cao nhất với P. maris. Tình trạng thiếu oxy và trao đổi chất ở ruột tôm và HP có thể thúc đẩy sự phát triển của Propionigenium trong quá trình tiến triển WFS.
Gần đây, axit succinic là một trong những chất chuyển hóa được phát hiện có liên quan tích cực với nhiều vi khuẩn gây bệnh như WFS và Vibrio. Axit succinic cũng là một nguồn cacbon cho Propionigenium tạo ra axit propionic. Ngoài ra, khẩu phần ăn bổ sung succinate ở tôm khỏe mạnh có thể gây ra sự thay đổi loại vi khuẩn đường ruột tương tự như ở tôm WFS (Huang và cộng sự, 2020). Các hướng nghiên cứu tiếp theo được đề xuất bao gồm 1) các thử nghiệm về khả năng axit propionic có thể gây ra sự hình thành bào tử và tổn thương HP ở tôm thẻ chân trắng, 2) nghiên cứu phân lập và nuôi Propionigenium từ tôm WFS để thử nghiệm sinh học với tôm nhiễm EHP và các loài xác định và 3) công việc dịch tễ học để xác định các yếu tố rủi ro (bao gồm sự hiện diện hay vắng mặt của các loài Propionigenium và Vibrio) liên quan đến các đợt bùng phát WFS.
5/ Kết luận
Nghiên cứu này cho thấy vi khuẩn ưu thế của chi Propionigenium và Vibrio cùng với vi bào tử trùng EHP là những nguyên nhân tiềm ẩn của một loại hội chứng phân trắng cụ thể được gọi là EHP-WFS ở tôm thẻ chân trắng nuôi. Trong một ao có biểu hiện ao EHP-WFS nghiêm trọng bao gồm cả việc tôm chết bất thường, tôm WFS có số bào tử EHP trong HP cao hơn so với tôm NG theo cả phân tích mô bệnh học và phân tử. Điều này phù hợp với tải lượng Propionigenium trong HP cao hơn và rối loạn hệ vi sinh vật ruột trong WFS so với tôm NG. Những kết quả này mở đường cho các nghiên cứu sâu hơn để kiểm tra giả thuyết rằng EHP và vi khuẩn trong chi Propionigenium và Vibrio tạo thành một bệnh lý sinh vật nhân chuẩn-nhân sơ có thể gây ra EHP-WFS.
Theo Natthinee Munkongwongsiri, Anuphap Prachumwat, Wiraya Eamsaard,
Kanokwan Lertsiri, Timothy W. Flegel, Grant D. Stentiford, Kallaya Sritunyalucksana,
Từ khóa: Hội chứng phân trắng (WFS), Penaeus vannamei, Enterocytozoon hepatopenaei (EHP), EHP-WFS, Propionigenium, Vibrio.
Biên dịch: Nguyễn Thị Quyên – Tôm Giống Gia Hóa Bình Minh
“Tôm Giống Gia Hóa – Chìa Khóa Thành Công”
Xem thêm:
- Chiết xuất trà xanh có thể chống lại WSSV và AHPND
- Liệu Những Đột Phá Về Gen Có Thổi Luồng Sinh Khí Mới Vào Nghề Nuôi Sú Và Tôm Càng Xanh Hay Không?
- Việt Nam Thử Sức Với Việc Nuôi Tôm Bằng Công Nghệ Semi Biofloc